§ 2. Основні компоненти еукаріотної клітини
Еукаріотні клітини (рис. 8 і 9) організовані значно складніше за прокаріотні. Дуже різноманітні вони і за своїми розмірами (від кількох мікрометрів до кількох сантиметрів), і формою, і структурними особливостями (рис. 10).
Рис. 8. Будова клітини еукаріотів. Узагальнена схема
Рис. 9. Будова клітини за даними електронної мікроскопії
Рис. 10. Різні еукаріотні клітини: 1 – епітеліальна; 2 – крові (е – еритроїїт, / – лейкоіїт); 3 – хряша; 4 – кістки; 5 – гладка м'язова; 6 – сполучної тканини; 7 – нервові клітини; 8 – поперечно-смугасте м'язове волокно
Проте загальна організація та наявність основних компонентів у всіх еукаріотних клітин однакові (рис. 11).
Рис. 11. Еукаріотна клітина (схема)
Плазмалемма (зовнішня клітинна мембрана).Основу плазмалеми, як і інших мембран у клітинах (наприклад, мітохондрій, пластид тощо), становить шар ліпідів, що має два ряди молекул (рис. 12). Оскільки молекули ліпідів полярні (один полюс вони гідрофілен, т. е. притягується водою, інший гідрофобен, т. е. відштовхується від води), те й розташовуються вони у порядку. Гідрофільні кінці молекул одного шару направлені у бік водного середовища – у цитоплазму клітини, а іншого шару – назовні від клітини – у бік міжклітинної речовини (у багатоклітинних) або водного середовища (у одноклітинних).
Рис. 12. Будова клітинної мембрани відповідно до рідинно-мозаїчної моделі. Білкії глікопротеїни занурені у подвійний шар ліпідних молекул, звернених своїми гідрофільними кінцями (кружки) назовні, а гідрофобними (хвилясті лінії) – углиб мембрани
Молекули білків мозаїчно вбудовані в бімолекулярний шар ліпідів. З зовнішнього боку тваринної клітини до ліпідів і молекул білків плазмалеми приєднуються молекули полісахаридів, утворюючи гліколіпіди та глікопротеїни.
Ця сукупність формує шар глікокаліксу.З ним пов'язана рецепторна функціяплазмалеми (див. нижче); також у ньому можуть накопичуватися різні речовини, що використовуються клітиною. Крім того, глікоколікс посилює механічну стійкість плазмалеми.
У клітинах рослин та грибів є ще клітинна стінка, що грає опорну та захисну роль. У рослин вона складається з целюлози, а у грибів – з хітину.
Зовнішня клітинна мембрана виконує ряд функцій, серед яких:
♦ механічна(опорна, формотворча);
♦ бар'єрно-транспортна(виборча проникність щодо різних речовин: надходження у клітину необхідних та виведення непотрібних та шкідливих);
♦ рецепторна(Визначення різних хімічних речовин, що опинилися в безпосередній близькості від клітини; сприйняття сигналів у вигляді гормонів; впізнавання «чужого» білка клітинами імунної системи і т. д.).
Обмін речовин між клітиною та навколишнім середовищем здійснюється різними способами – пасивними та активними.
Молекули води та різних іонів пасивно (за рахунок дифузії, осмосу), без витрати клітиною енергії, надходять через особливі пори – це пасивний транспорт.Макромолекули, такі як білки, полісахариди, навіть цілі клітини, надходять шляхом фагоцитозі піноцитозіз витратою енергії – активний транспорт.
Шляхом фагоцитозу поглинаються цілі клітини або великі частинки (наприклад, згадайте про харчування у амеб або фагоцитоз захисними клітинами крові бактерій). При піноцитоз відбувається поглинання дрібних частинок або крапель рідкої речовини. Загальним для обох процесів є те, що поглинаються речовини оточуються зовнішньою мембраною, що вп'ячується, з утворенням вакуолі, яка потім переміщається в глиб цитоплазми клітини.
Екзоцитоз є процес (будучи також активним транспортом), протилежний у напрямку фагоцитозу і піноцитозу (рис. 13). З його допомогою можуть виводитись неперетравлені залишки їжі у найпростіших або утворені в секреторній клітині біологічно активні речовини.
цитоплазма.Цитоплазма – це вміст клітини, обмежений плазмалемою, крім ядра. У її складі виділяють основна речовина (гіалоплазму), органоїдиі включення.
Гіалоплазма- в'язка рідина, здатна перебувати в стані або золя(рідком), або гелю(студноподібний).
При необхідності цитоплазма здатна оборотно переходити з одного стану до іншого. Наприклад, при амебоїдному русі (згадайте розділ «Найпростіші» з курсу зоології) в ході утворення ложноніжки відбуваються швидкі переходи цитоплазми з гелю в золь і навпаки. Це зумовлено наявністю в цитоплазмі великої кількості ниткоподібних молекул із білка актину.Коли вони, з'єднуючись один з одним, утворюють тривимірну мережу, цитоплазма перебуває у стані гелю, а коли мережа розпадається – може золя.
У гіалоплазмі містяться різні речовини – ферменти, білки, вуглеводи, жири та інші, органічні та мінеральні. Тут здійснюються різні хімічні процеси – розщеплення речовин, їх синтез та модифікації (зміни).
Органоїди.Це постійні компоненти клітини з певною будовою та функціями, що перебувають у її цитоплазмі. Надалі йтиметься про органоїди загального призначення,властивих будь-яким типам клітин усіх еукаріотів. Із ними пов'язане забезпечення життєдіяльності останніх. Органоїди спеціального призначеннязустрічаються лише у клітинах певного (вузькоспеціалізованого) типу – наприклад, міофібрили у м'язових клітинах.
Органоїди загального призначення мають однакову будову незалежно від того, яким клітинам та яких організмів вони належать. Але серед них виділяють групи з мембранним (ендоплазматична мережа, апарат Гольджі, мітохондрії, пластиди, лізосоми, вакуолі),а також немембранним ( рибосоми, клітинний центр)будовою.
Ендоплазматична мережа (ЕПС).ЕПС складається з мембран і є складно розгалуженою системою канальців і цистерн, що пронизує всю цитоплазму клітини (рис. 14). Розрізняють два види ЕПС - шорсткуі гладку.До мембран шорсткої (з боку цитоплазми) прикріплюються рибосоми, а на гладкій їх немає.
Рис. 14. Ендоплазматична мережа
Ендоплазматична мережа виконує в еукаріотній клітині низку найважливіших функцій:
♦ що розмежовує(Поділ внутрішнього обсягу клітини на різні реакційні простори);
♦ участь у синтезі органічних речовин(на мембранах шорсткої ЕПС розташовуються рибосоми, але в гладкої – ферментні комплекси, які забезпечують синтез ліпідів, вуглеводів тощо. буд.);
♦ участь у формуванні елементів апарату Гольджі, лізосом;
♦ транспорт речовин.
Апарат Гольджі.Апарат Гольджі (АГ) є системою цистерн(плоських вакуолей) та бульбашок(Везикул), розташовану в безпосередній близькості від ядра клітини, які утворюються за рахунок ЕПС в результаті відділення невеликих її фрагментів (рис. 15). При злитті цих фрагментів і виникають нові цистерни апарату Гольджі, при цьому з ЕПС транспортуються різні речовини, які беруть участь у складанні складних органічних сполук (білки + вуглеводи, білки + ліпіди тощо), що виводяться за допомогою АГ за межі клітини. Ці біологічно активні речовини або виводяться із клітини (за допомогою секреторних вакуолей шляхом екзоцитозу), або входять до складу лізосом (див. нижче), що утворюються за рахунок АГ.
Рис. 15. Апарат Гольджі:
Апарат Гольджі виконує такі функції:
♦ синтезбіологічно активних речовин, що виробляються клітиною;
♦ секрецію (виведення з клітини) різних речовин(гормонів, ферментів, речовин, з яких будується клітинна стінка, тощо);
♦ участь у освіті лізосом.
Мітохондрії.Мітохондрії є у всіх типів еукаріотних клітин (рис. 16). Вони мають вигляд або округлих тілець, або паличок, рідше – ниток. Їхні розміри коливаються від 1 до 7 мкм. Число мітохондрій у клітці становить від кількох сотень до десятків тисяч (у великих найпростіших).
Рис. 16. Мітохондрії. Вгорі – мітохондрії (a) у сечових каналах, видимі у світловому мікроскопі. Внизу – тривимірна модель організації мітохондрії: 1 – кристи; 2 – зовнішня мембрана; 3 – внутрішня мембрана; 4 – матрикс
Мітохондрія утворена двома мембранами – зовнішньоїі внутрішньої,між якими розташовано міжмембранний простір.Внутрішня мембрана утворює безліч вп'ячувань – христ, що є або пластини, або трубочки. Така її організація забезпечує величезну площу внутрішньої мембрани. На ній розташовуються ферменти, що забезпечують перетворення енергії, що міститься в органічних речовинах (вуглеводах, ліпідах), в енергію АТФ, необхідну для життєдіяльності клітини. Отже, функція мітохондрій – участь у енергетичнихклітинних процесах. Саме тому велика кількість мітохондрій притаманна, наприклад, м'язовим клітинам, які виконують велику роботу.
Пластиди.У рослинних клітинах виявляються особливі органоїди - пластиди, що мають частіше веретеноподібну або округлу форму, іноді складнішу. Розрізняють три види пластид – хлоропласти (рис. 17), хромопласти та лейкопласти.
Хлоропластивідрізняються зеленим кольором, який обумовлений пігментом. хлорофілом,що забезпечує процес фотосинтезу,тобто синтезу органічних речовин з води (Н 2 О) та вуглекислого газу (СО 2) з використанням енергії сонячного світла. Хлоропласти містяться переважно у клітинах листя (у вищих рослин). Вони сформовані двома паралельно розташованими один одному мембранами, що оточують вміст хлоропластів. строму.Внутрішня мембрана утворює численні сплощені мішечки. тилакоїди,які складені в стопки (на зразок стопки монет) – грани –і лежать у стромі. Саме в тилакоїдах і міститься хлорофіл.
Хромопластивизначають жовтий, помаранчевий та червоний колір багатьох квіток та плодів, у клітинах яких присутні у великій кількості. Основними пігментами у складі є каротини.Функціональне призначення хромопластів полягає у колірному залученні тварин, що забезпечують запилення квіток та поширення насіння.
Рис. 17. Пластиди: а – хлоропласти у клітинах листа елодеї, видимі у світловому мікроскопі; б – схема внутрішньої будови хлоропласту з гранами, що являють собою стоси плоских мішечків, розташованих перпендикулярно поверхні хлоропласту; в – більш докладна схема, на якій видно анастомозуючі трубочки, що з'єднують окремі камери фан
Лейкопласти– це безбарвні пластиди, що містяться у клітинах підземних частин рослин (наприклад, у бульбах картоплі), насіння та серцевини стебел. У лейкопластах, головним чином, відбувається утворення з глюкози крохмалю і накопичення його в органах рослин, що запасають.
Пластиди одного виду можуть перетворюватися на інший. Наприклад, при осінній зміні кольору листя хлоропласти перетворюються на хромопласти.
Лізосоми.Ці органоїди мають вигляд бульбашок, оточених мембраною, діаметром до 2 мкм. Вони містять кілька десятків ферментів, що розщеплюють білки, нуклеїнові кислоти, полісахариди та ліпіди. Функція лізосом – участь у процесах внутрішньоклітинного розщеплення складних органічних сполук (наприклад, харчових речовин або речовин, що «відпрацювали» клітинних компонентів). Лізосоми зливаються з фагоцитарними (або піноцитарними) вакуолями, утворюючи травну вакуоль.
Утворення лізосом відбувається за рахунок відпочкування від цистерн апарату Гольджі.
Рибосоми.Рибосоми (рис. 18) присутні в клітинах як еукаріотів, так і прокаріотів, оскільки виконують важливу функцію в біосинтезі білків(Див. розділ 5). У кожній клітині є десятки, сотні тисяч (до кількох мільйонів) цих дрібних округлих органоїдів.
Рис. 18. Схема будови рибосоми, яка сиділа на мембрані ендоплазматичної мережі: 1 – мала суб'єдиниця; 2 – тРНК; 3 – аміноацил-тРНК; 4 – амінокислота; 5 – велика субодиниця; 6 – мембрана ендоплазматичної мережі; 7 – синтезований поліпептидний ланцюг
Рибосома складається з двох нерівних субодиниць (частин). Вони утворюються окремо і поєднуються, «охоплюючи» інформаційну РНК, у процесі синтезу білкової молекули. До складу рибосом входять різні білки та рибосомальні РНК.
Клітинні включення.Так називаються непостійні компоненти у клітині, присутні у основному речовині цитоплазми як зерен, гранул чи крапельок. Включення можуть бути оточені мембраною або не оточуються нею.
У функціональному відношенні виділяють три види включень: запасні поживні речовини(крохмаль, глікоген, жири, білки), секреторні включення(речовини, характерні для залізистих клітин, що продукуються ними, - гормони залоз внутрішньої секреції тощо) включення спеціального призначення(У вузькоспеціалізованих клітинах, наприклад гемоглобін в еритроцитах).
§ 3. Організація клітинного ядра. Хромосоми
Клітинне ядро (див. рис. 8 і 9) має найважливіше значення у життєдіяльності клітини, оскільки служить сховищем спадкової інформації, що міститься в хромосомах (див. нижче).
Ядро обмежене ядерною оболонкою, що відокремлює його вміст (Каріоплазму)від цитоплазми. Оболонка і двох мембран, розділених проміжком. Обидві вони пронизані численними порами, завдяки яким можливий обмін речовин між ядром і цитоплазмою. У ядрі клітини у більшості еукаріотів знаходиться від 1 до 7 ядерців.З ними пов'язані процеси синтезу РНК та тРНК.
Основні компоненти ядра хромосоми,утворені з молекули ДНК та різних білків. У світловому мікроскопі вони добре помітні лише під час клітинного поділу (Мітоз, мейоз).У клітині, що не ділиться, хромосоми мають вигляд довгих тонких ниток, розподілених по всьому об'єму ядра.
Під час поділу клітин хромосомні нитки утворюють щільні спіралі, унаслідок чого стають видимими (за допомогою звичайного мікроскопа) у формі паличок, «шпильок». Весь обсяг генетичної інформації розподілено між хромосомами ядра. У процесі вивчення були виявлені такі закономірності:
♦ в ядрах соматичних клітин (тобто клітин тіла, нестатевих) у всіх особин одного виду міститься однакова кількість хромосом, що становлять набір хромосом(Рис. 19);
Рис. 19. Хромосоми різних видів рослин та тварин, зображені в одному масштабі: 1,2 – амеба; 3,4 – діатомові водорості; 5–8, 18,19 – зелені водорості; 9 – мухомор; 10 – липа; 11–12 – дрозофіла; 13 - сьомга; 14 – скерда (родина складноцвітих); 15 – рослина із сімейства ароїдних; 16 - метелик-чубатка; 17 – комаха із сімейства саранчових; 20 - клоп-водомірка; 21 - квітковий клоп; 22 – земноводна амбістома; 23 - алое (сімейство лілейних)
♦ для кожного виду характерний свій хромосомний набір за їх кількістю (наприклад, у людини 46 хромосом, мушка дрозофіли – 8, аскарида – 4, річковий рак – 196, конь – 66, кукурудза – 104);
♦ хромосоми в ядрах соматичних клітин можуть бути згруповані парами, що отримали назву гомологічне хромосомна підставі їх подібності (за будовою та функціями);
♦ в ядрах статевих клітин (гамет) з кожної пари гомологічних хромосом міститься лише одна, тобто загальний набір хромосом вдвічі менший, ніж у соматичних клітинах;
♦ одинарний набір хромосом у статевих клітинах називається гаплоїднимі позначається буквою n, а соматичних – диплоїдним(2n).
З викладеного ясно, кожна пара гомологічних хромосом утворена об'єднанням батьківських і материнських хромосом при заплідненні, т. е. злиття статевих клітин (гамет). І навпаки, при утворенні статевих клітин із кожної пари гомологічних хромосом у гамету потрапляє лише одна.
Хромосоми різнихгомологічні пари відрізняються за розмірами і формою (рис. 20 і 21).
Рис. 20. Будова та типи хромосом: а – зовнішній вигляд 1 – цетроміру; 2 – коротке плече; 3 – довге плече); внутрішня структура тієї ж хромосоми (1 – центромір; молекули ДНК); в – типи хромосом (1 – одноплеча; різноплеча; 3 – рівноплеча: X – плече, V – центроміру)
Рис. 21. Хромосома складається з ДНК та білків. Молекула ДНК реплікується. Дві ідентичні подвійні спіралі ДНК залишаються з'єднаними в області іентромера. Ці копії перетворюються на окремі хромосоми пізніше, під час поділу клітини
У тілі хромосом виділяють первинну перетяжку (звану центромірою),до якої прикріплюються нитки веретена поділу.Вона ділить хромосому на два плеча.Хромосоми можуть бути рівноплечими, різноплечими та одноплечими.
Розділ 5. Обмін речовин
§ 1. Метаболізм як єдність асиміляції та дисиміляції
Всі клітини та живі організми – це відкриті системи, тобто вони перебувають у стані постійного обміну енергій та речовин із навколишнім середовищем. Є відкриті системи й у неживої природі, та їх існування якісно відрізняється від живих організмів. Розглянемо такий приклад: шматок самородної сірки, що горить, знаходиться в стані обміну з навколишнім середовищем. При його горінні поглинається О 2, а виділяються SO 2 та енергія (у вигляді тепла). Однак при цьому шмат сірки як фізичне тіло руйнується, втрачає свою первинну структуру.
Для живих організмів обмін з навколишнім середовищем виявляється умовою збереження, підтримки їх структурної організації шляхом самооновлення всіх речовин і компонентів, з яких вони складаються.
Обмін речовин (метаболізм) - сукупність процесів, що протікають в живих організмах (споживання, перетворення, накопичення і виділення речовин і енергії), що забезпечують їх життєдіяльність, розвиток, зростання, відтворення. У процесі обміну речовин відбувається розщеплення та синтез молекул, що входять до складу клітин; оновлення клітинних структур та міжклітинної речовини.
В основі метаболізму лежать взаємопов'язані процеси асиміляції(анаболізм) та дисиміляції(Катаболізм). При асиміляції (пластичний обмін) відбувається синтез складних речовин із простих. Саме завдяки цьому створюються всі органічні речовини у клітині, необхідні для побудови її структурних компонентів, ферментних систем тощо. Асиміляція завжди здійснюється із витратою енергії.
У ході дисиміляції (енергетичний обмін) складні органічні речовини розщеплюються до простих чи неорганічних. При цьому виділяється енергія, яка витрачається клітиною на виконання різних процесів, що забезпечують її життєдіяльність (синтез та транспорт речовин, механічну роботу тощо).
Всі живі організми можуть бути поділені на дві групи: автотрофиі гетеротрофи,які відрізняються джерелами енергії та необхідних речовин для забезпечення своєї життєдіяльності.
Автотрофи– організми, що синтезують із неорганічних речовин органічні сполуки з використанням енергії сонячного світла (як фототрофи- рослини, ціанобактерії або енергії, одержуваної при окисленні мінеральних (неорганічних) речовин (таких, як хемотрофи– серобактерії, залізобактерії та ін.). Отже, вони здатні самостійно створювати необхідні речовини для своєї життєдіяльності.
§ 2. Дисиміляція у анаеробних та аеробних організмів
Організми можуть бути поділені на дві групи та за характером дисиміляції – аеробиі анаероби.Аероби (від грец. біс– повітря) потребують вільного кисню для життєдіяльності. У анаеробів (грец. ац- Негативна частка) в ньому немає необхідності. У них дисиміляція здійснюється шляхом бродіння - безкисневого, ферментативного розщеплення органічної речовини з утворенням більш простих органічних речовин і виділенням енергії. Наприклад:
♦ молочнокисле бродіння:
C 6 H 12 O 6 + 2H 3 PO 4 + 2АДФ → 2Ф Н + 2АТФ + 2H 2 O;
♦ спиртове бродіння:
C 6 H 12 O 6 + 2Ф Н + 2АДФ → 2C 2 H 5 OH + 2CO 2 + 2АТФ + 2H 2 O.
Речовини, що утворюються при бродінні, є органічними і, отже, містять ще багато енергії.
Рис. 22. Взаємини асиміляції та дисиміляції у автотрофних та гетеротрофних організмів
У аеробних організмів у процесі дихання в мітохондріях відбувається повне розщеплення органічних речовин (при використанні Про 2) до бідних енергією кінцевих продуктів СО 2 іН 2 Про і вивільняється значно більша кількість енергії:
З 6 Н 12 0 6 (глюкоза) + 0 2 > 6С0 2 + 6Н 2 0 + енергія (за рахунок якої синтезується 38 молекул АТФ).
Розглянемо як узагальнених схем метаболізм у автотрофних і гетеротрофних аеробних організмів (рис. 22).
Асиміляція.Її найважливіші процеси – фотосинтезі біосинтез білків.
Глава 1
ОСНОВИ КЛІТИННОЇ ФІЗІОЛОГІЇ
І. Дудель
Плазматична мембрана . Тварини клітини обмежені плазматичною мембраною (рис. 1.1). На її будові, дуже подібній до будови багатьох внутрішньоклітинних мембран, ми зупинимося дещо докладніше. Основний матрикс мембрани складається з ліпідів,головним чином фосфатидил-холіну. Ці ліпіди складаються з головної гідрофільної групи, до якої приєднані довгі гідрофобні вуглеводневі ланцюги. У воді такі ліпіди спонтанно формують двошарову плівку товщиною 4-5 нм, у якій гідрофільні групи звернені до водного середовища, а гідрофобні вуглеводневі ланцюги розташовуються у два ряди, утворюючи безводну ліпідну фазу. Клітинні мембрани є ліпідними бислоями саме такого типу і містять гліколіпіди, холестерол і фосфоліпіди (рис. 1.2). Гідрофільна частина гліколіпідів утворена олігосахаридами. Гліколіпіди завжди розташовуються на зовнішній поверхні плазматичної мембрани, причому олігосахаридна частина молекули орієнтована подібно до волоска, зануреного в навколишнє середовище. Розкидані серед фосфоліпідів у майже рівній кількості молекули холестеролу стабілізують мембрану. Розподіл різних ліпідів у внутрішньому та зовнішньому шарах мембрани неоднаковий, і навіть у межах одного шару є ділянки, у яких концентруються окремі види ліпідів. Такий нерівномірний розподіл,
Рис. 1.1. Схематичний малюнок клітини, на якому зображені найважливіші органели
мабуть, має якесь, поки що неясне, функціональне значення.
Головними функціональними елементами, зануреними у порівняно інертний ліпідний матрикс мембрани, є білки(Рис. 1.2). Білок по масі становить від 25 до 75% у різних мембранах, але оскільки білкові молекули набагато більші, ніж ліпідні, 50% по масі еквівалентні співвідношенню: 1 молекула білка на 50 молекул ліпіду. Одні білки пронизують мембрану від її зовнішньої до внутрішньої поверхні, інші ж закріплені в якомусь одному шарі. Білкові молекули зазвичай орієнтовані так, що їх гідрофобні групи занурені у ліпідну мембрану, а полярні гідрофільні групи на поверхні мембрани занурені у водну фазу. Багато білків зовнішньої поверхні мембрани є глікопротеїни; їх гідрофільні сахаридні групи звернені у позаклітинне середовище.
Мембранні системи внутрішньоклітинних органел .
Приблизно половина об'єму клітини зайнята органелами ізольованими від цитозолю мембранами. Загальна поверхня мембран внутрішньоклітинних органел принаймні у 10 разів перевищує поверхню плазматичної мембрани. Найбільш поширена мембранна система - ендоплазматичний ретикулум,являє собою мережу

Рис. 1.2.Схематичне зображення плазматичної мембрани. Білки занурені у фосфоліпідний бішар, причому деякі з них пронизують бішар, тоді як інші тільки заякорені на зовнішньому або внутрішньому шарі.
сильно звивистих трубочок або мішковидних витягнутих структур; великі ділянки ендоплазматичного ретикулуму усіяні рибосомами; такий ретикулум отримав назву гранулярного, або шорсткого (рис. 1.1). Апарат Гольджітакож складається з мембранзв'язних ламел, від яких відриваються бульбашки або везикули (рис. 1.1). Лізосоми та пероксисоми– це невеликі спеціалізовані везикули. У всіх цих різноманітних органелах мембрана і простір, що охоплюється нею, містять специфічні набори ферментів; всередині органел накопичуються особливі продукти обміну, використовувані реалізації різних функцій органел .
Ядроі мітохондріївідрізняються тим, що кожна з цих органел оточена двома мембранами. Ядро є відповідальним за кінетичний контроль метаболізму; складчаста внутрішня мембрана мітохондрії – місце окисного метаболізму; тут за рахунок окислення пірувату або жирних кислот синтезується високоенергетична сполука аденозинтрифосфат (АТФ, або АТР).
Цитоскелет . Цитоплазму, що оточує органели, аж ніяк не можна вважати аморфною; її пронизує мережа цитоскелета. Цитоскелет складається з мікротрубочок, актинових філаментів та проміжних філаментів (рис. 1.1). Мікротрубочкимають зовнішній діаметр близько 25 нм; вони утворюються, як і звичайний полімер, внаслідок збирання молекул білка тубуліна. Актинові філаменти-скорочувальні волокна, що розташовуються в примембранному шарі та у всій клітині – переважно беруть участь у процесах, пов'язаних з рухом. Проміжні філаментискладаються з блоків різного хімічного складу у різних типах клітин; вони утворюють різноманітні зв'язки між згаданими вище двома іншими елементами цитоскелету. Органели та плазматична мембрана також пов'язані з цитоскелетом, який не тільки забезпечує підтримання форми клітини та положення органел у ній, але й обумовлює зміну форми клітини та її рухливість.
Цитозоль . Близько половини обсягу клітини зайнято цитозолем. Оскільки він приблизно на 20% (за масою) складається з білка, це швидше за гель, ніж водний розчин. Малі молекули, у тому числі органічні та неорганічні іони,розчинені у водній фазі. Між клітиною та навколишнім середовищем (позаклітинним простором) відбувається обмін іонів; ці обмінні процеси будуть розглянуті у наступному розділі. Концентрація іонів у позаклітинному просторі підтримується зі значною точністю постійному рівні; внутрішньоклітинна концентрація кожного з іонів також має специфічний рівень, який відрізняється від такого поза клітиною (табл. 1.1). Найбільш поширеним катіоном у позаклітинному середовищі є Na + у клітині його концентрація більш ніж у 10 разів нижча. Навпаки, всередині клітини найбільш висока концентрація К + поза клітиною вона нижче більш ніж на порядок. Найбільший градієнт між позаклітинною та внутрішньоклітинною концентраціями існує для Са 2+ концентрація вільних іонів якого всередині клітини принаймні у 10000 разів нижча, ніж поза нею. Не всі іони розчинені в цитозолі, їх частина адсорбована на білках або депонована в органелах. Наприклад, у випадку Са 2+ пов'язані іони набагато численніші, ніж вільні. Більшість білків цитозолю – ферменти, з участю яких здійснюється безліч процесів проміжного метаболізму: гліколіз і глюконеогенез, синтез чи руйнація амінокислот, синтез білка на рибосомах (рис. 1.1). У цитозолі містяться також краплі ліпідів та гранули глікогену, що служать резервами важливих молекул.
Таблиця 1.1.Внутрішньо- та позаклітинні концентрації іонів у м'язових клітинах гомойотермних тварин. А – -«високомолекулярні клітинні аніони»
|
внутрішньоклітинна концентрація |
Позаклітинна концентрація |
||
|
Na + |
12 ммоль/л |
Na + |
145 ммоль/л |
|
155 ммоль/л |
До + |
4 ммоль/л |
|
|
Са 2+ |
10 -7 10 -8 ммоль/л |
Са 2+ |
2 ммоль/л |
|
З l - |
4 ммоль/л |
З 1 - |
120 ммоль/л |
|
HCO 3 – |
8 ммоль/л |
HCO 3 – |
27 ммоль/л |
|
A – |
155 ммоль/л |
Інші катіони |
5 ммоль/л |
|
Потенціал спокою -90 мВ |
|||
1.2. Обмін речовинами між клітиною та навколишнім середовищем
Ми коротко описали будову клітини, щоб використати цей опис для розгляду основ клітинної фізіології. Клітину в жодному разі не можна вважати статичним утворенням, оскільки тут відбувається постійний обмін речовин між різними внутрішньоклітинними компартментами, а також між компартментами і навколишнім середовищем. Структури клітини знаходяться в динамічній рівновазі, і взаємодії клітин між собою та із зовнішнім середовищем є необхідною умовою для підтримання життя функціонуючого організму. У цьому розділі ми розглянемо фундаментальні механізми такого обміну. У наступних розділах ці механізми будуть розглянуті у додатку до нервової клітини та її функцій;
однак ті самі механізми лежать в основі функціонування всіх інших органів.
Дифузія.Найпростішим процесом переміщення речовини є дифузія. У розчинах (або газах) атоми та молекули переміщуються вільно, а різниця концентрацій урівноважується завдяки дифузії. Розглянемо два обсяги, заповнені рідиною чи газом (рис. 1.3), у яких речовини мають концентраціїз 1 і з 2 та розділені шаром з площею поверхні А та товщиною d. Потік речовини m за час t описується першим законом дифузії Фіка:
dm/ dt= DA/ d ( C 1 -З 2) =DA/ dD C(1)
де D-коефіцієнт дифузії, постійний для даної речовини, розчинника та температури. У більш загальному вигляді для різниці концентрацій dc на відстані dx
dm/dt= -D A dc/dx,(2)
потік через переріз А пропорційний градієнту концентрації dc/dx . Знак «мінус» виникає у рівнянні тому, що зміна концентрації у напрямку х-негативна.
Дифузія – це найважливіший процес, завдяки якому більшість молекул у водних розчинах переміщається на невеликі відстані. Це відноситься і до їхнього руху в клітині остільки, оскільки дифузії не перешкоджають мембрани. Безліч речовин можуть вільно дифундувати через ліпідні мембрани, особливо вода і розчинені гази, такі, як 2 і 2 . Жиророзчинні

Рис. 1.3.Кількісна схема дифузії. Два простори розділені шаром завтовшкиdі площею А.С;-висока концентрація частинок у лівій частині обсягу, С:,-низька концентрація частинок у правій частини, рожева поверхня-Концентраційний градієнт в дифузійному шарі. Дифузійний потік dm/dt-див. рівняння (1)
речовини також добре дифундують через мембрани; це стосується і полярних молекул досить малого розміру, таких, як етанол та сечовина, тоді як цукри проходять через ліпідний шар насилу. Натомість ліпідні шари практично непроникні для заряджених молекул, у тому числі навіть для неорганічних іонів. Для неелектролітів рівняння дифузії (1) зазвичай перетворюють, з'єднуючи характеристики мембрани і дифузної речовини в один параметр-проникність (Р):
dm/dt=P AD с.(3)
На рис. 1.4 порівнюються проникності (Р) ліпідної мембрани для різних молекул
Дифузія через мембранні пори . Плазматична мембрана (та інші клітинні мембрани) проникні не тільки для речовин, що дифундують через ліпідний шар, але і для багатьох іонів, цукрів, амінокислот та нуклеотидів. Ці речовини долають мембрану через пори, утворені транспортними білками,зануреними у мембрану. Усередині таких білків є заповнений водою канал діаметром менше 1 нм, через який можуть дифундувати малі молекули. Вони рухаються по градієнту концентрації, і якщо вони несуть заряд, їх рух каналами регулюється також мембранним потенціалом. Мембранні канали мають відносну вибірковість.

Рис. 1.4.Проникність штучних ліпідних бішарів для різних речовин
По відношенню до типу молекул, які можуть через них проходити, існують, наприклад, калієві, натрієві та кальцієві канали, кожен з яких непроникний практично для будь-якого іона, крім специфічного. Така вибірковістьобумовлена зарядом або структурою місць зв'язування у стінках каналу, що полегшує транспорт специфічної молекули та запобігає проникненню через канал інших речовин (мал. 1,5, А) .
За поведінкою мембранних іонних каналівлегко спостерігати, оскільки струм, що виникає при русі іонів, можна виміряти, причому навіть для одиночного каналу. Показано, що канали спонтанно та з високою частотою змінюють свій стан від відкритого до закритого. Для калієвого каналу характерні імпульси струму амплітудою близько 2 пА (2 10 -12 А) і тривалістю кілька мілісекунд (див. рис. 2.12, стор 37) [З]. За цей період через нього проходять десятки тисяч іонів. Перехід білків з однієї конформації до іншої вивчають методами рентгенівської дифракції, мессбауерівської спектроскопії та ядерно-магнітного резонансу (ЯМР). Білки, таким чином, є дуже динамічними рухомими структурами, а канал, що проходить через білок, не просто жорстка, наповнена водою трубка (мал. 1.5, А), але молекулярних груп і зарядів, що швидко рухаються. Ця динамічна характеристика каналу відображається в енергетичному профілі каналу,показаному на рис. 1.5, Б. Тут по осі абсцис представлена довжина каналу від зовнішнього розчину з концентрацією іонів 0 і потенціалом 0 до внутрішнього розчину з концентрацією 1 і потенціалом Е. По осі ординат

Рис. 1.5.А. Схема білка, що утворює калієвий канал, занурений у ліпідний бислой плазматичної мембрани. Чотири негативні заряди зафіксовані на «стінці» каналу. Б. Схематичний енергетичний профіль каналу показаного на рис. А. По осі ординат відкладені величини кінетичної енергії, необхідних проходження каналу; по осі абсцис -відстань між внутрішньою і зовнішньою поверхнями мембрани. Енергетичні мінімуми відповідають місцям зв'язування позитивно заряджених іонів із фіксованими негативними зарядами у стінці каналу. Енергетичні максимуми відповідають перешкод дифузії в каналі. Передбачається, що конформація канального білка спонтанно осцилює; варіанти енергетичних профілів зображені суцільними та штриховими лініями; ці осциляції значною мірою полегшують зв'язування іонів при подоланні енергетичного бар'єру (за змінами)
представлені рівні енергії іона у місцях зв'язування каналу; пік на графіку представляє бар'єр проникності, який енергія іона повинна перевершувати для проникнення через канал, а «провал» графіка – порівняно стабільний стан (зв'язування). Незважаючи на перешкоду у вигляді енергетичного піку, він може проникнути через канал, якщо енергетичний профіль спонтанно циклічно змінюється; іон, таким чином, може раптово виявитися "по той бік" енергетичного піку і може продовжити рух у клітину. Залежно від заряду, розмірів та ступеня гідратації іона та його здатності зв'язуватися зі структурами стінок каналу енергетичний профіль каналу варіює для різних іонів, чим може пояснюватися вибірковість окремих типів каналів.
Дифузійна рівновага іонів . Дифузія різних іонів через мембранні канали мала б призвести до усунення відмінностей у концентраціях між поза- та внутрішньоклітинними середовищами. Як, проте, видно із табл. 1.1, такі відмінності зберігаються, отже, має бути якесь рівновагаміж дифузією та іншими процесами транспорту через мембрану Наступні два розділи стосуються тих шляхів, за допомогою яких така рівновага встановлюється. У разі іонів на дифузійну рівновагу впливає їхній заряд. Дифузія незаряджених молекул забезпечується різницею концентрацій dc , і коли концентрації зрівнюються, власне транспорт припиняється. На заряджені частинки впливає електричне поле. Наприклад, коли іон калію виходить із клітини за градієнтом своєї концентрації, він несе один позитивний заряд. Таким чином, внутрішньоклітинне середовище стає більш негативно зарядженим, внаслідок чого на мембрані виникає різниця потенціалів. Внутрішньоклітинний негативний заряд перешкоджає виходу з клітини нових іонів калію, а ті іони, які все ж таки вийдуть з клітини, ще більше збільшать заряд на мембрані. Потік іонів калію припиняється, коли дія електричного поля компенсує дифузійний тиск унаслідок різниці концентрацій. Іони продовжують проходити через мембрану, але у рівних кількостях в обох напрямках. Отже, для даної різниці концентрацій іонів на мембрані існує рівноважний потенціалЕ ion при якому потік іонів через мембрану припиняється. Рівноважний потенціал може бути легко визначений за допомогою рівняння Нернста:
Еion= RT/ zF* lnC out/ C in(4)
де R -газова постійна, Т-абсолютна температура, z - валентність іона (негативна для аніонів) C out - позаклітинна концентрація іону, C in -Внутрішньоклітинна концентрація іона, F число Фарадея. Якщо підставити в рівняння константи, то за температури тіла (Т = 310 К) рівноважний потенціал для іонів калію Е K дорівнює:
Ек= –61 мB log /(5)
Якщо [ K + out ]/[ K + in ] = 39, як це випливає з табл. 1.1, тоді
Ек = -61 м B log 39 = -97 мВ.
Дійсно, було виявлено, що всі клітини мають мембранний потенціал;у м'язових клітинах ссавців рівень становить близько -90 мВ. В залежності від умов та відносних концентрацій іонів клітини можуть мати мембранний потенціал у межах від -40 до -120мВ. Для клітини у наведеному вище прикладі (табл. 1.1) потенціал спокою,рівний приблизно -90 мВ показує, що потоки іонів калію через мембранні канали знаходяться приблизно в рівновазі. Це не дивно, оскільки в мембрані, що лежить, відкритий стан калієвих каналів найбільш ймовірно, тобто. мембрана найбільш проникна для іонів калію. Мембранний потенціал, однак, визначається потоками та інших іонів.
Легкість, з якою незаряджені частинки можуть дифундувати через мембрану, кількісно представлена у рівнянні (3). Проникність для заряджених частинок описується дещо складнішим рівнянням:
P= m RT/ dF(6)
де m-рухливість іона в мембрані, d -Товщина мембрани, a R , Т та F -відомі термодинамічні постійні. Визначені таким чином значення проникності для різних іонів можуть бути використані для розрахунку мембранного потенціалу Em , коли іони калію, натрію та хлору проходять через мембрану одночасно (з проникністю Р K , Р Na та P Cl відповідно). При цьому передбачається, що потенціал падає в мембрані рівномірно, тому напруженість поля постійна. У цьому випадку застосовується рівняння Голдмана, або рівняння постійного поля :
Еm= R T/F * ln(P K + P Na + P Cl )/ (P K + P Na + P Cl )(7)
Для більшості клітинних мембран Р K приблизно 30 разів вище, ніж Р Na (Див. також розд. 1.3). Відносна величина P Cl сильно варіює; для багатьох мембран P Cl мала в порівнянні з Р K однак для інших (наприклад, у скелетних м'язах) P Cl значно вище, ніж Р K.
Активний транспорт, натрієвий насос . У попередньому розділі описані пасивна дифузія іонів і мембранний потенціал, що утворюється внаслідок цього, при заданих внутрішньо- і позаклітинних концентраціях іонів. Однак у результаті цього процесу концентрація іонів усередині клітини стабілізується не автоматично, оскільки мембранний
потенціал дещо більш негативний, ніж E K , і набагато – порівняно з Е Na (близько +60 мВ). Завдяки дифузії внутрішньоклітинні концентрації іонів, принаймні калію та натрію, повинні зрівнюватися з позаклітинними. Стабільність градієнта іонів досягається за допомогою активного транспорту: мембранні білки переносять іони через мембрану проти електричного та (або) концентраційного градієнтів, споживаючи для цього метаболічну енергію. Найважливіший процес активного транспорту – це робота Na/K -насоса, що існує практично у всіх клітинах;
насос викачує іони натрію із клітини, одночасно накачуючи іони калію всередину клітини. Таким чином забезпечується низька внутрішньоклітинна концентрація іонів натрію та висока калію (табл. 1.1). Градієнт концентрації іонів натрію на мембрані має специфічні функції, пов'язані з передачею інформації у вигляді електричних імпульсів (див. Розд. 2.2), а також з підтримкою інших активних транспортних механізмів та регулювання об'єму клітини (див. нижче). Тому не дивно, що більше 1/3 енергії, що споживається клітиною, витрачається на Na/K-насос, а в деяких найбільш активних клітинах на його роботу витрачається до 70% енергії.
Na/K-транспортний білок є АТФазу. На внутрішній поверхні мембрани вона розщеплює АТФ на АДФ та фосфат (рис. 1.6). На транспортування трьох іонів натрію з клітини одночасно двох іонів калію в клітину використовується енергія однієї молекули АТФ, т. е. сумарно за цикл з клітини видаляється один позитивний заряд. Таким чином, Na/K-насос є електрогенним(Створює електричний струм через мембрану), що призводить до збільшення електронегативності мембранного потенціалу приблизно на 10 мВ. Транспортний білок виконує цю операцію з високою швидкістю: від 150 до 600 іонів натрію за секунду. Амінокислотна послідовність транспортного білка відома, проте ще зрозумілий механізм цього складного обмінного транспорту. Цей процес описують за допомогою енергетичних профілів перенесення білками іонів натрію або калію (рис. 1.5,5). За характером зміни цих профілів, пов'язаних із постійними змінами конформації транспортного білка (процес, що вимагає витрати енергії), можна судити про стехіометрію обміну: два іони калію обмінюються на три іони натрію.
Na/K-насос, як і ізольована Na + /К + -залежна мембранна АТФаза, специфічно інгібується серцевим глікозидом уабаїном (строфантіном). Оскільки робота Na/K–насоса є багатоступінчастою хімічною реакцією, вона, подібно до всіх хімічних реакцій, значною мірою залежить від температури, що

Рис. 1.6.Схема Na/K-насоса-АТФази (зануреної в ліпідний бислой плазматичної мембрани), яка за один цикл виносить з клітини три іони Na + проти градієнтів потенціалу та концентрації і приносить у клітину два іони К + . У процесі цього одна молекула АТФ розщеплюється на АДФ і фосфат. На схемі АТФаза показана як димер, що складається з великої (функціональної) та малої субодиниць; у мембрані вона існує як тетрамер, утворений двома великими та двома малими субодиницями
продемонстровано на мал. 1.7. Тут потік іонів натрію з м'язових клітин показаний щодо часу; практично це еквівалентно потоку іонів натрію, опосередкованому роботою Na/K-насоса, тому що пасивний потік іонів натрію проти градієнтів концентрації та потенціалу вкрай малий. Якщо препарат охолодити приблизно на 18°С, то потік іонів натрію з клітини швидко зменшиться у 15 разів, а відразу після нагрівання відновиться до початкового рівня. Таке зменшення потоку іонів натрію з клітини в кілька разів більше, ніж те, що відповідало б температурній залежності процесу дифузії або простої хімічної реакції. Подібний ефект спостерігається, коли запас метаболічної енергії виснажується через отруєння динітрофенолом (ДНФ) (рис. 1.7,5). Отже, потік іонів натрію із клітини забезпечується енергозалежною реакцією – активним насосом. Іншою характеристикою насоса поряд із значною температурною та енергетичною залежністю є наявність рівня насичення (як і у всіх інших хімічних реакцій); це означає, що швидкість роботи насоса не може зростати нескінченно при підвищенні концентрації іонів, що транспортуються (рис. 1.8). На відміну від цього потік пасивно дифузної речовини зростає пропорційно різниці концентрацій відповідно до закону дифузії (рівняння 1 і 2).
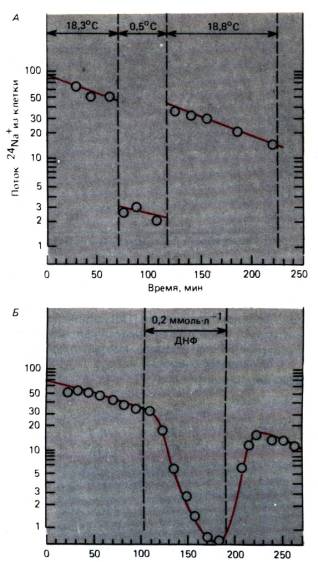
Рис. 1.7. А, Б.Активний транспорт Na +. Вісь ординат:потік радіоактивного 24 Na + клітини (імп./хв). Ось абсцис:час початку експерименту. А.Клітину охолоджено з 18,3 °С до 0,5 °С; потік Na + із клітини у цей період загальмований. Б.Пригнічення потоку Na + з клітини динітрофенолом (ДНФ) у концентрації 0,2 ммоль/л (із змінами)
Крім Na/K-насоса плазматична мембрана містить принаймні ще один насос – кальцієвий;це насос відкачує іони кальцію (Са 2+) з клітини та бере участь у підтримці їх внутрішньоклітинної концентрації на вкрай низькому рівні (табл. 1.1). Кальцієвий насос присутній з дуже високою густиною в саркоплазматичному ретикулумі м'язових клітин, які накопичують іони кальцію внаслідок розщеплення молекул АТФ (див. гл. 4).
Вплив Na/K-насоса на мембранний потенціал та об'єм клітини . На рис. 1.9 показані різні компоненти мембранного струму та наведені внутрішньоклітинні концентрації іонів, які

Рис. 1.8.Співвідношення між швидкістю транспорту молекул та їх концентрацією (у місці входу в канал або в місці зв'язування насоса) при дифузії через канал або насосному транспорті. Останній при високих концентраціях насичується (максимальна швидкість, Vmax ) значення по осі абсцис, що відповідає половині максимальної швидкості насоса ( Vmax /2), є рівноважною концентрацією До m

Рис. 1.9.Схема, що показує концентрації Na + , K+ та Cl – всередині та поза клітини та шляхи проникнення цих іонів через клітинну мембрану (через специфічні іонні канали або за допомогою Na/K–насоса. При даних концентраційних градієнтах рівноважні потенціали E Na , E K та E С l – рівні вказаним, мембранний потенціал Em = – 90 мВ
забезпечують їхнє існування. Через калієві канали спостерігається струм іонів калію, що виходить, так як мембранний потенціал дещо більш електропозитивний, ніж рівноважний потенціал для іонів калію. Загальна провідність натрієвих каналів набагато нижча, ніж калієвих, тобто. натрієві канали відкриті набагато рідше, ніж калієві при потенціалі спокою; однак у клітину входить приблизно стільки ж іонів натрію, скільки виходить з неї іонів калію, тому що для дифузії іонів натрію в клітину необхідні великі градієнти концентрації та потенціалу. Na/K-насос забезпечує ідеальну компенсацію пасивних дифузійних струмів, тому що переносить іони натрію з клітини а іони калію-в неї. Таким чином, насос є електрогенним за рахунок різниці в числі перенесених у клітину та з клітини зарядів, що при нормальній швидкості його роботи створює мембранний потенціал приблизно на 10 мВ більш електронегативний, ніж якщо він утворювався тільки за рахунок пасивних потоків іонів (див. рівняння 7). В результаті мембранний потенціал наближається до калієвого рівноважного потенціалу, що зменшує витік іонів калію. Активність Na/K-насоса регулюється внутрішньоклітинною концентрацією іонів натрію. Швидкість роботи насоса уповільнюється при зниженні концентрації іонів натрію, що підлягають виведенню з клітини (рис. 1.8), так що робота насоса та потік іонів натрію всередину клітини врівноважують один одного, підтримуючи внутрішньоклітинну концентрацію натрію іонів на рівні приблизно 10 ммоль/л.
Щоб підтримувати рівновагу між насосним та пасивними мембранними струмами, необхідно набагато більше молекул Na/K–насоса, ніж канальних білків для іонів калію та натрію. При відкритому стані каналу через нього проходять десятки тисяч іонів за кілька мілісекунд (див. вище), а оскільки канал зазвичай відкривається кілька разів на секунду, за цей час через нього проходить більше 10 5 іонів. Одиночний насосний білок переміщає кілька сотень іонів натрію в секунду, отже, плазматична мембрана повинна містити приблизно 1000 разів більше насосних молекул, ніж канальних. Вимірювання канальних струмів у спокої показало наявність у середньому одного калієвого та одного натрієвого відкритого каналу на 1 мкм 2 мембрани; з цього випливає, що у тому просторі має бути близько 1000 молекул Na/K–насоса, тобто. відстань між ними становить середньому 34 нм; діаметр насосного білка як канального становить 8-10 нм. Таким чином, мембрана досить щільно насичена насосними молекулами.
Той факт, що потік іонів натрію всередину клітини, а іонів калію з клітини компенсується роботою насоса, є й інше слідство, що полягає у збереженні стабільного осмотичного тиску постійного обсягу.Усередині клітини існує висока концентрація великих аніонів, головним чином білків (А – у табл. 1.1), які не здатні проникати через мембрану (або проникають через неї дуже повільно) і тому є фіксованим компонентом усередині клітини. Щоб урівноважити заряд цих аніонів, потрібна рівна кількість катіонів. Завдяки дії Na/K-насоса цими катіонами в основному виявляються іони калію. Істотне підвищення внутрішньоклітинної концентрації іонів могло б відбуватися лише при зростанні концентрації аніонів внаслідок потоку С1 – за градієнтом концентрації у клітину (табл. 1.1), але мембранний потенціал протидіє цьому. Вхідний струм Cl – спостерігається тільки доти, доки не буде досягнуто рівноважного потенціалу для іонів хлору; це спостерігається, коли градієнт іонів хлору практично протилежний градієнту іонів калію, тому що іони хлору заряджені негативно (рівняння 4). Таким чином, встановлюється низька внутрішньоклітинна концентрація іонів хлору, що відповідає низькій позаклітинній концентрації іонів калію. Результатом є обмеження загальної кількості іонів у клітині. Якщо мембранний потенціал падає при блокаді Na/K-насоса, наприклад, при аноксії, то рівноважний потенціал для іонів хлору знижується, а внутрішньоклітинна концентрація іонів хлору відповідно підвищується. Відновлюючи рівновагу зарядів, іони калію також входять у клітину; сумарна концентрація іонів у клітині зростає, що підвищує осмотичний тиск; це змушує воду надходити у клітку. Клітина набухає. Таке набухання спостерігається in vivo в умовах нестачі енергії.
Концентраційний градієнт Na + як рушійна сила мембранного транспорту . Значення Na/K-насоса для клітини не обмежується стабілізацією нормальних градієнтів К+ та Na+ на мембрані. Енергія, запасена в мембранному градієнті Na + часто використовується для забезпечення мембранного транспорту інших речовин. Наприклад, на рис. 1.10 показано «симпорт» Na + та молекули цукру в клітину. Мембранний транспортний білок переносить молекулу цукру в клітину навіть проти градієнта концентрації, в той же час Na + рухається по градієнту концентрації та потенціалу, забезпечуючи енергію для транспорту цукрів.Такий транспорт цукру повністю залежить від існування високого градієнта. Na + ; якщо внутрішньоклітинна концентрація Na + значно зростає, то транспорт цукрів припиняється. Для різних c ахарів існують різні симпортні системи. Транспорт амінокислоту клітку подібний до транспорту c ахарів, показаним на рис. 1.10; він також забезпечується градієнтом Na + , існує принаймні п'ять різних систем симпорта, кожна з яких спеціалізована для будь-якої однієї групи родинних амінокислот.
Крім симпортних систем існують також "антипортні".Одна з них, наприклад, за один цикл переносить один іон кальцію з клітини в обмін на три вхідні іони натрію (рис. 1.10). Енергія для транспорту Са 2+утворюється за рахунок входу трьох іонів натрію по градієнту концентрації та потенціалу. Цієї енергії достатньо (при потенціалі спокою) для підтримки високого градієнта іонів кальцію (від менше 10 -7 моль/л усередині клітини до приблизно 2 ммоль/л поза клітиною).
Ендо- та екзоцитоз . Для деяких речовин, що надходять у клітину або мають бути виведені

Рис. 1.10.Білки, занурені в ліпідний біс мембрани, опосередковують симпорт глюкози і Na + в клітину, а також Ca 2+ / Na + -Антипорт, в якому рушійною силою є градієнт Na + на клітинній мембрані
з неї транспортні канали відсутні; до таких речовин відносяться, наприклад, білки та холестерол. Вони можуть проходити через плазматичну мембрану везикулах,або бульбашках,за допомогою ендо- та екзоцитозу. На рис. 111 показані основні механізми цих процесів. При екзоцитозі певні органели (див. нижче) формують везикули, заповнені речовиною, яку необхідно вивести з клітини, наприклад, гормонами або ферментами позаклітинної дії. Коли такі везикули досягають плазматичної мембрани, їхня ліпідна мембрана зливається з нею, даючи таким чином можливість вмісту вийти у зовнішнє середовище. При протилежному процесі-ендоцитозі-плазматична мембрана інвагінує, утворюючи ямку, яка потім поглиблюється і замикається, формуючи внутрішньоклітинну везикулу, заповнену позаклітинною рідиною та деякими макромолекулами. Щоб забезпечити це злиття мембран і замикання везикули, скорочувальні елементи цитоскелета діють разом із самими мембранами (див. нижче). При ендоцитозі який завжди відбувається просто захоплення позаклітинної середовища в клітину. У клітинній мембрані містяться часто організовані спеціалізовані групи специфічні рецептори до макромолекул, таким, як інсулін або антигени. Після того, як ці макромолекули зв'яжуться зі своїми рецепторами, в оточуючому ділянці мембрани відбувається ендоцитоз, і макромолекула вибірково транспортується в клітину (рис. 1.12, Б).
Ендо- та екзоцитоз відбуваються в клітинах безперервно. Кількість мембранного матеріалу, що робить оборот, значно; протягом 1 години макрофаг поглинає у вигляді везикул подвійну площу поверхні своєї цитоплазматичної мембрани. У більшості клітин оборот мембранного матеріалу відбувається не настільки інтенсивно, але все ж таки повинен бути значним.

Рис. 1.11.Екзоцитоз та ендоцитоз. Вгорі:внутрішньоклітинна везикула зливається з ліпідним бислоем плазматичної мембрани та відкривається у позаклітинний простір. Цей процес називається екзоцитозом. Внизу:плазматична мембрана інвагінує на невеликій ділянці та відшнуровує везикулу, заповнену позаклітинним матеріалом. Цей процес називають ендоцитозом
1.3. Перенесення речовин усередині клітини
Ендо- та екзоцитоз-це не тільки процеси транспорту речовин через клітинну мембрану, а й процеси обміну мембран – структурних компонентів самої клітини. Предметом розгляду в цьому розділі є інші подібні транспортні процеси в клітині та її органелах.

Рис. 1.12. А-В.Схема процесів, що включають екзо- та ендоцитоз. А.Білок, синтезований у гранулярному ендоплазматичному ретикулумі, транспортується за допомогою апарату Гольджі до плазматичної мембрани, де секретується екзоцитозом. Б.Холестерол, пов'язаний з частинками ЛНП (ліпопротеїну низької щільності), приєднується до плазматичної мембрани, індукує утворення ендоцитозної бульбашки в цій ділянці мембрани та транспортується до лізосом, де вивільняється. Ст.Позаклітинний матеріал, захоплений у процесі ендоцитозу (на малюнку праворуч),транспортується через клітину у везикулах, або бульбашках, та виділяється за допомогою екзоцитозу (на малюнку ліворуч)
Дифузія . Природно, що у цитозоле різниця концентрацій усувається з допомогою дифузії; те саме справедливо і для рідин, укладених в органелах. Через високу концентрацію розчиненого білка дифузія тут протікає набагато повільніше, ніж у воді. Ліпідні мембрани – навколо клітини та у складі органел – це двомірні рідини, у яких відбувається дифузія. Ліпіди в мембранному бішарі дифундують у межах власного шару, рідко переходячи з одного в інший. Занурені у яких білки також досить рухливі; вони обертаються навколо осі, перпендикулярної до мембрани, або латерально дифундують з дуже різними константами дифузії, в 2-10000 разів повільніше, ніж фосфоліпіди. Так, якщо деякі білки переміщуються в ліпідному шарі вільно і з такою самою швидкістю, як самі ліпідні молекули, інші заякорені, тобто. досить міцно пов'язані з цитоскелетом. Існують «постійні» агрегати специфічних білків у мембрані, наприклад пре-і постсинаптичні структури нервових клітин. білки, що вільно переміщаються, можна продемонструвати шляхом зв'язування їх з флуоресцентними барвниками, світіння яких індукують, висвітлюючи короткочасними спалахами невелику ділянку мембрани. Такі експерименти показують, що менш ніж за 1 хв білки, що зв'язалися з барвником, рівномірно розподіляються по мембрані на відстані до 10 мкм.
Активний транспорт у мембранах органел .
Процеси активного транспорту, які відіграють життєво важливу роль у функціонуванні плазматичної мембрани, протікають також і всередині клітини – у мембранах органел. Специфічний вміст різних органел створюється частково завдяки внутрішньому синтезу, а частково – за рахунок активного транспорту з цитозолю. Одним із прикладів останнього є згадуваний вище Са 2+ насос в саркоплазматичному ретикулумі м'язових клітин. Особливо цікаво, що у разі синтезу АТФ у мітохондріях діє принцип, зворотний тому, що має місце у АТФазних насосах плазматичної мембрани (рис. 1.6). При синтезі АТФ окисний метаболізм призводить до утворення крутого градієнта H+ на внутрішніх мембранах. Цей градієнт є рушійною силою для процесу, зворотного насосному циклу активного транспорту молекул: іони Н + рухаються через мембрану по градієнту, а енергія, що вивільняється в результаті цього, забезпечує синтез АТФ з АДФ і фосфату. АТФ, що утворилася, у свою чергу забезпечує енергією клітину, у тому числі і для активного транспорту.
Транспорт у везикулах . У клітці є велика кількість органел і пов'язаних із нею везикул (рис. 1.1). Ці органели, а везикули особливо, знаходяться у постійному русі, транспортуючи свій вміст до інших органелів або до плазматичної мембрани. Везикули можуть також мігрувати з клітинної мембрани до органел, як при ендоцитозі.
Процес секреції білкапредставлений на рис. 1.12, А.Білок синтезується поблизу ядра клітини на рибосомах, пов'язаних з ендоплазматичним ретикулумом (так званий гранулярний або шорсткий, ендоплазматичний ретикулум); потрапивши в ендоплазматичний ретикулум, білок упаковується в транспортні везикули, які відокремлюються від органели та мігрують до апарату Гольджі. Тут вони зливаються з цистернами апарату Гольджі, де білок модифікується (тобто перетворюється на глікопротеїн). На кінцях цистерн везикули знову відокремлюються. Несучі модифікований білок секреторні везикули рухаються до плазматичної мембрани та виділяють вміст шляхом екзоцитозу.
Інший приклад транспортного шляху у клітині показаний на рис. 1.12, Б; це-поглинання холестеролу клітиною. Холестерол, що транспортується в крові, пов'язаний в основному з білками, наприклад частинками «ліпопротеїну низької щільності»(ЛНП). Такі частинки приєднуються до специфічних, що містять рецептори до ЛНП ділянок мембрани, де відбувається ендоцитоз і ЛНП переноситься всередину клітини в «окаймлених» везикулах. Ці везикули зливаються, утворюючи ендосоми і втрачаючи в ході цього процесу «окаймлення». Ендосоми у свою чергу зливаються з первинними лізосомами, що містять переважно гідролітичні ферменти, і утворюють вторинні, більші лізосоми. Вони холестерол вивільняється з частинок ЛНП і дифундує в цитозоль, де стає доступним, наприклад, для синтезу ліпідних мембран. Від ендосом також відокремлюються везикули, що не містять ЛНП, які особливим шляхом рухаються до плазматичної мембрани та зливаються з нею, повертаючи мембранний матеріал та, ймовірно, рецептори до ЛНП. З моменту зв'язування частки ЛНП із мембраною до вивільнення холестеролу з вторинної лізосоми проходить 10-15 хв. Порушення у зв'язуванні та поглинанні ЛНП, тобто у постачанні клітини холестеролом, відіграють вирішальну роль у розвитку серйозного та широко поширеного захворювання – атеросклерозу («затвердіння» артерій).
Існує безліч інших транспортних шляхів, подібних до показаних на рис. 1.11 і 1.12, А, за допомогою яких рухаються в клітці специфічні везикули. Невідомо, як саме вони пересуваються, але в цей процес, ймовірно, залучені елементи цитоскелету. Везикули можуть ковзати мікротрубочками, в цьому випадку енергія для руху, мабуть, забезпечується пов'язаним з везикулами білком - АТФазою (див. нижче). Залишається зовсім незрозумілим, як безліч різних везикул, рухаючись одна за одною у всіх напрямках, потрапляють за призначенням. Вони, очевидно, мають бути «помічені» таким чином, щоб це розпізнавалася транспортною системою та перетворювалося на цілеспрямований рух.
Транспорт шляхом утворення та руйнування органел . Досі ми розглядали ендо- та екзоцитоз як процеси транспортування вмісту везикул. Існує й інший аспект цих процесів, що полягає в тому, що спрямоване видалення плазматичної мембрани на одній ділянці клітинної поверхні шляхом ендоцитозу і, навпаки, додавання її іншим шляхом екзоцитозу переміщує значні ділянки мембрани (рис. 1.12.Д), даючи клітині можливість, наприклад , сформувати виріст чи рухатися.
Подібні перебудови типові також і для цитоскелета, особливо для мікрофіламентів та мікротрубочок (рис. 1.1). Мікрофіламентискладаються в першу чергу з білка F-актину,який здатний до збирання волокнисті пучки в результаті полімеризації мономеру з цитозолю. Пучки поляризовані, тобто часто наростають тільки з одного кінця, акумулюючи нові молекули актину, тоді як інший кінець інертний або тут відбувається розбирання. За рахунок такого поляризованого зростання мікрофіламенти ефективно переміщуються і може змінюватися структура мережі. Перехід актину з деполімеризованого стану (золя) до організованого (гель) може відбуватися дуже швидко під дією інших білків або змін концентрації іонів (див. нижче). Існують також білки, які спричиняють руйнування актинових філаментів з утворенням коротких фрагментів. Тонкі вирости багатьох клітин – філоподії – містять центральний пучок актину (рис. 1.1), і різні рухи філоподії, ймовірно, зумовлені переходами актину: полімеризація – деполімеризація.
Мікротрубочкитакож часто зазнають таких переміщень. Механізм цих переміщень подібний - полімеризація тубуліна з цитозолю таким чином, що один з кінців мікротрубочки росте, тоді як інший або не змінюється, або відбувається розбирання. Так мікротрубочка шляхом відповідного додавання або усунення матеріалу може переміщуватися цитозолю.
Активні рухи цитоскелету . Зміни цитосклетних структур можуть відбуватися в результаті активних рухів, так і перебудов, описаних вище. У багатьох випадках рух мікротрубочок та актинових філаментів обумовлено скоротливими білками, які пов'язують філаменти або трубочки та можуть переміщувати їх відносно один одного. Білки міозин та дінеїнприсутні у цитозолі всіх клітин у порівняно високих концентраціях; вони є тими елементами, які перетворюють енергію в рух у спеціалізованих клітинах (м'язових) та органелах (віях). У м'язових клітинах міозин утворює товсті філаменти, орієнтовані паралельно до актинових філаментів. Молекула міозину своєю «головкою» приєднується до актинового філаменту та, використовуючи енергію АТФ, зміщує міозин вздовж молекули актину.Потім міозин від'єднується від актину. Сукупність безлічі таких циклів з'єднання-роз'єднання призводить до макроскопічного скорочення м'язових волокон(Гл. 4). Дінеїн грає аналогічну роль у переміщенні мікротрубочок при роботі вій (рис. 1.1). У цитоплазмі неспеціалізованих клітин міозин і дінеїн утворюють не правильні волокна, а здебільшого невеликі групи молекул. Навіть у вигляді таких малих агрегатів вони можуть переміщати актинові філаменти або мікротрубочки. Рис. 1.13 ілюструє цей процес, коли до двох актинових філаментів, поляризованих у різних напрямках, також приєднані протилежно поляризовані молекули міозину. Головні групи міозину згинаються до хвоста молекули, витрачаючи при цьому АТФ, а два актинові філаменти зміщуються в протилежному напрямку, після чого міозин від'єднується від них. Переміщення такого роду, у ході яких енергія АТФ перетворюється на механічну роботу, можуть змінювати форму цитоскелета і, отже, клітини, і навіть забезпечувати транспорт пов'язаних із цитоскелетом органел.
Процеси внутрішньоклітинного транспорту найяскравіше можуть бути продемонстровані на аксоні нервової клітини. Аксонний транспортрозглядається тут докладно, щоб проілюструвати події, які, ймовірно, подібним чином відбуваються у більшості клітин. Аксон, діаметр якого становить лише кілька мікронів, може досягати довжини одного метра і більше, і рух білків шляхом дифузії від ядра до дистального кінця аксона зайняло б роки. Давно відомо, що коли будь-якої з ділянок аксона піддається констрикції, частина аксона, розташована проксимальніше, розширюється. Це виглядає так, ніби в аксон блокований відцентровий потік. Такий потік-швидкий аксонний транспорт можебути продемонстрований рухом радіоактивних маркерів, як у експерименті, показаному на рис. 1.14. Лейцин, мічений радіоактивною міткою, ін'єктували в ганглій дорсального корінця, і потім з 2-ї по 10-ю годину вимірювали радіоактивність у сідничному нерві на відстані 166 мм від тіл нейронів. За 10 годин пік радіоактивності у місці ін'єкції змінювався незначно. Але хвиля радіоактивності поширювалася по аксону з постійною швидкістю близько 34 мм за 2 год, або 410 мм/сут. Показано, що у всіх нейронах гомойотермних тварин швидкий аксонний транспорт здійснюється з такою ж швидкістю, причому відчутних відмінностей між тонкими, безмієліновими волокнами та найбільш товстими аксонами, а також між моторними та сенсорними волокнами не спостерігається. Тип радіоактивного маркера також впливає швидкість швидкого аксонного транспорту; маркерами можуть бути різноманітні радіоактивні

Рис. 1.13.Нем'язовий міозиновий комплекс за певної орієнтації може зв'язуватися з актиновими філаментами різної полярності та, використовуючи енергію АТФ, зміщувати їх відносно один одного
молекули, такі як різні амінокислоти, що включаються в білки тіла нейрона. Якщо проаналізувати периферичну частину нерва, щоб визначити природу переносників транспортованої радіоактивності сюди, то такі переносники виявляються головним чином у фракції білків, але також у складі медіаторів і вільних амінокислот. Знаючи, що властивості цих речовин різні і особливо різні розміри їх молекул, постійну швидкість транспорту ми можемо пояснювати лише загальним всім них транспортним механізмом.
Описаний вище швидкий аксонний транспортє антероградним,тобто спрямованим від тіла клітини. Показано, що деякі речовини рухаються від периферії до тіла клітини за допомогою ретроградний транспорт.Наприклад, ацетилхолінестераза транспортується в цьому напрямку зі швидкістю в 2 рази меншою, ніж швидкість швидкого аксонного транспорту. Маркер, який часто використовується в нейроанатомії пероксидаза хрону – також переміщається ретроградним транспортом. Ретроградний транспорт, ймовірно, відіграє важливу роль у регуляції синтезу білка в тілі клітини. Через кілька днів після перерізання аксона у тілі клітини спостерігається хроматоліз, що свідчить про порушення білкового синтезу. Час, потрібний для хроматолізу, корелює з тривалістю ретроградного транспорту від місця перерізання аксона до тіла клітини. Такий результат передбачає пояснення цього порушення – порушується передача з периферії «сигнального речовини», регулюючого білковий синтез. Очевидно, що основними «засобами пересування», що використовуються для швидкого аксонного

Рис. 1.14.Досвід, що демонструє швидкий аксонний транспорт у сенсорних волокнах сідничного нерва кішки. Мічений тритієм лейцин вводять у ганглій дорсального корінця та вимірюють радіоактивність у ганглії та сенсорних волокнах через 2, 4, 6, 8 та 10 год після введення (Нижня частина малюнка).за осі абсцисвідкладено відстань від ганглія до ділянок сідничного нерва, де вимірюють. По осі ординат тільки для верхньої та нижньої кривої в логарифмічному масштабі відкладено радіоактивність (імп./хв). «Хвиля» підвищеної радіоактивності (стрілки)рухається зі швидкістю 410мм/сут (по )
транспорту, є везикули (бульбашки) та органели,такі як мітохондрії, що містять речовини, які потрібно транспортувати. Переміщення найбільших везикул або мітохондрії можна спостерігати за допомогою мікроскопа in vivo . Такі частинки здійснюють короткі швидкі рухи в одному з напрямків, зупиняються, часто рухаються трохи назад або убік, знову зупиняються, а потім ривок в основному напрямку. 410 мм/сут відповідають середньої швидкості антероградного руху приблизно 5 мкм/с; швидкість ж кожного окремого руху повинна бути, отже, значно вищою, а якщо врахувати розміри органел, філаментів та мікротрубочок, то ці рухи дійсно дуже швидкі. Швидкий аксонний транспорт потребує значної концентрації АТФ. Такі отрути, як колхіцин, що руйнує мікротрубочки, також блокують швидкий аксонний транспорт. З цього випливає, що в аналізованому нами транспортному процесі везикули та органели рухаються вздовж мікротрубочок та актинових філаментів; цей рух забезпечується малими агрегатами молекул динеїну та міозину, що діють, як показано на рис. 1.13 з використанням енергії АТФ.
Швидкий аксонний транспорт може брати участь і в патологічні процеси.Деякі нейротропні віруси (наприклад, віруси герпесу або поліомієліту) проникають в аксон на периферії і рухаються за допомогою ретроградного транспорту до тіла нейрона, де розмножуються і мають свою токсичну дію. Токсин правця-білок, який продукується бактеріями, що потрапляють в організм при пошкодженнях шкіри, захоплюється нервовими закінченнями та транспортується до тіла нейрона, де він викликає характерні м'язові спазми. Відомі випадки токсичного на сам аксонний транспорт, наприклад вплив промисловим розчинником акриламідом. Крім того, вважають, що патогенез авітамінозу "бері-бері" та алкогольної полінейропатії включає порушення швидкого аксонного транспорту.
Крім швидкого аксонного транспорту в клітці існує досить інтенсивний повільний аксонний транспорт.Тубулін рухається по аксону зі швидкістю близько 1 мм/сут, а актин швидше - до 5мм/сут. З цими компонентами цитоскелета мігрують інші білки; наприклад, ферменти, очевидно, пов'язані з актином чи тубуліном. Швидкості переміщення тубуліна і актину приблизно узгоджуються зі швидкістю зростання, виявленою для механізму, описаного раніше, коли молекули включаються до активного кінця мікротрубочки або мікрофіламенту. Отже, цей механізм може бути основою повільного аксонного транспорту. Швидкість повільного аксонного транспорту приблизно відповідає також швидкості зростання аксона, що, мабуть, вказує на обмеження, що накладаються структурою цитоскелету на другий процес.
Завершуючи цей розділ, слід наголосити, що клітини в жодному разі не є статичними структурами, якими вони здаються, наприклад, на електронно-мікроскопічних фотографіях. Плазматична мембранаі особливо органели знаходяться в постійному швидкому русі та постійній перебудові;Тільки тому вони здатні працювати. Далі це не прості камери, в яких протікають хімічні реакції, а високоорганізовані конгломерати мембран та волокон,в яких реакції протікають оптимально організованої послідовності.
1.4. Регулювання клітинних функцій
Підтримка індивідуальної клітини як функціональної одиниці переважно регулюється ядром; вивчення таких регуляторних механізмів – предмет клітинної біології та біохімії. Разом з тим клітини повинні модифікувати свої функції відповідно до умов навколишнього середовища та потреб інших клітин організму, тобто вони служать об'єктами функціонального регулювання. Нижче ми коротко розглянемо, яким чином ці регуляторні впливи діють на плазматичну мембрану і як вони сягають внутрішньоклітинних органел.
Регуляторні дії на клітинну мембрану
Мембранний потенціал . У багатьох випадках регулювання клітинних функцій здійснюється шляхом зміни мембранного потенціалу. Можливі локальні зміни потенціалу, коли: 1) струм із сусідньої ділянки клітини або генерований іншою клітиною протікає через мембрану; 2) змінюється позаклітинна концентрація іонів (часто [К+] out ); 3) відкриваються мембранні іонні канали. Зміни мембранного потенціалу можуть проводити конформацію мембранних білків, змушуючи, зокрема, відкриватися чи закриватися канали. Як описано вище, функціонування деяких мембранних насосів залежить від мембранного потенціалу. Нервові клітини спеціалізовані сприймати зміни мембранного потенціалу як інформацію, яка має перероблятися та передаватись (див. гл. 2).
Позаклітинні регуляторні речовини . Найбільш важливим регуляторним механізмом за участю позаклітинних речовин є їхня взаємодія зі специфічними рецепторами на плазматичній мембрані або всередині клітини. До таких речовин відносяться синаптичні медіатори, які передають інформацію між нервовими клітинами, локальні агенти та речовини, що циркулюють у крові та досягають всіх клітин організму, наприклад гормони та антигени. Синаптичні медіаториє невеликими молекулами, що виділяються з нервових закінчень в області синапсу;
коли вони досягають плазматичної мембрани сусідньої, постсинаптичної клітини вони запускають електричні сигнали або інші регуляторні механізми. Це питання докладно розглянуто у гол. 3.
Локальні хімічні агенти часто виділяються спеціалізованими клітинами. Вони вільно дифундують у позаклітинному просторі, проте їхня дія обмежена невеликою групою клітин внаслідок швидкого руйнування цих речовин, або спонтанного, або під дією ферментів. Одним із прикладів виділення таких агентів є вивільнення гістамінуопасистими клітинами при пошкодженні або імунної реакції. Гістамін викликає розслаблення гладком'язових клітин судин, збільшує проникність судинного ендотелію та стимулює сенсорні нервові закінчення, що опосередковують відчуття сверблячки. Інші локальні хімічні агенти виділяються багатьма іншими клітинами. Типовими локальними агентами є простагландини,складові групу приблизно 20 похідних жирних кислот. Вони виділяються безперервно з поширених клітин, але діють лише локально, оскільки швидко руйнуються фосфоліпазами мембрани. Різні простагландини мають широкий спектр дії: вони можуть запускати скорочення гладких клітин, викликати агрегацію кров'яних пластинок (тромбоцитів) або пригнічувати розвиток жовтого тіла в яєчниках.
Інші локальні агенти служать факторами зростання.Найбільш відомий фактор росту нервів (ФРН) для симпатичних нейронів, який необхідний для зростання та виживання цих нейронів у ході розвитку in vivo або у клітинній культурі. Очевидно, клітини-мішені для цього класу нейронів виділяють ФРН і тим самим забезпечують правильну іннервацію. При формуванні органів клітин часто необхідно «знаходити шлях» до клітин-мішеней, які можуть розташовуватися на значних відстанях. Відповідно, має існувати безліч спеціалізованих факторів зростання, подібних до ФРН.
Гормони та антигени переносяться кров'ю до всіх клітин. Антигени викликають імунну відповідь клітин, що несуть специфічні антитіла. Однак антигени, як правило, є чужорідними речовинами, які не утворюються в реагує організмі (детальніше див. гл. 18). Деякі гормони, такі, як інсулін або тироксин, впливають на клітини, що відносяться до найрізноманітніших типів, тоді як інші, наприклад, статеві гормони, – тільки на клітини певного типу. Гормони це пептиди, дія яких запускається зв'язуванням їх з рецептором на клітинній мембрані, або стероїди і тироксин, які дифундують через ліпідну мембрану і зв'язуються з внутрішньоклітинними рецепторами. Стероїдні гормони зв'язуються з хроматином ядер, у результаті запускається транскрипція певних генів. Продукуються в результаті білки викликають зміну клітинних функцій, в чому полягає специфічна дія гормонів. Питання, пов'язані з виділенням та дією гормонів, докладно розглянуті у гол. 17.
Внутрішньоклітинна комунікація за участю других посередників
Регуляторні функції, описані вище, включають на клітинну мембрану. Інформація, отримана мембраною клітини, часто повинна викликати реакцію органел і переноситься до них різними речовинами, відомими як другі посередники (на відміну від перших, що надходять до клітин від зовнішніх джерел). Вивчення других посередників розвивається швидко, немає гарантії, що нинішній рівень розуміння проблеми виявиться досить повним. Тут ми торкнемося трьох добре вивчених посередників: Са 2+, цАМФ та інозитолтрифосфату.
Кальцій.Найпростіший внутрішньоклітинний посередник - це іон Са 2+. Його вільна концентрація в клітці, що покоїться, дуже низька і становить 10 _ -8 -10 -7 моль/л. Він може проникати в клітину через специфічні мембранні канали, коли вони знаходяться у відкритому стані, наприклад, при змінах мембранного потенціалу (див. гл. 2). Підвищення концентрації Са 2+, що виникає в результаті, запускає важливі реакції в клітині, такі, як скорочення міофібрил, яке є основою м'язового скорочення (див. гл. 4), або виділення везикул, що містять медіатори, з нервових закінчень (див. гл. 3) . Обидві реакції вимагають концентрації Са 2+ приблизно приблизно 10 -5 моль/л. Са 2+ , що має регуляторну дію, може вивільнятися також і з внутрішньоклітинних депо, таких як ендоплазматичний ретикулум. Вивільнення Са 2+ із депо потребує участі інших посередників (див., наприклад, рис. 1.16).
Циклічний аденозинмонофосфат, цАМФ. Останнім часом доведено, що циклічний аденозинмонофосфат (цАМФ), похідне основного джерела енергії в організмі-АТФ, є важливим другим посередником. Складний ланцюг реакцій, показаний на рис. 1.15, починається з рецептора R s на зовнішній поверхні плазматичної мембрани, який може бути місцем специфічного зв'язування для різних медіаторів та гормонів. Після зв'язування зі специфічною «стимулюючою» молекулою R s змінює свою конформацію; ці зміни впливають на білок G s на внутрішній поверхні мембрани таким чином, що стає можливою активація останнього внутрішньоклітинним гуанозинтрифосфатом (ГТФ). Активований білок G s , своєю чергою, стимулює фермент на внутрішній поверхні мембрани–аденилатциклазу (АЦ), яка каталізує утворення цАМФ з АТФ. Водорозчинний цАМФ є посередником, що передає ефект.

Рис. 1.15.Ланцюг реакцій за участю внутрішньоклітинного посередника цАМФ (циклічного аденозинмонофосфату). Збудливий або гальмівний зовнішні сигнали активують мембранні рецептори R s або Ri . Ці рецептори регулюють процес зв'язування G -білків з внутрішньоклітинним ГТФ (гуанозинтрифосфатом), стимулюючи або інгібуючи цим внутрішньоклітинну аденілатциклазу (АЦ). Підсилюючий фермент АЦ перетворює аденозинтрифосфат (АТФ) на цАМФ, який потім за участю фосфодіестераеи розщеплюється до АМФ. Вільний цАМФ дифундує в клітину та активує аденілаткіназу (А-кіназу), вивільняючи її каталітичну субодиницю С, яка каталізує фосфорилювання внутрішньоклітинних білків, тобто. формує кінцевий ефект позаклітинного стимулу. На схемі показані також фармакологічні препарати та токсини, які запускають (+) або інгібують (-) деякі реакції (за змінами)
стимуляції позаклітинного рецептора R s до внутрішніх структур клітини.
Паралельно зі стимуляторним ланцюгом реакцій за участю R s можливе зв'язування гальмівних медіаторів та гормонів з відповідним рецептором R i який знову-таки через ГТФ-активований білок G , інгібує АЦ і, таким чином, продукцію ЦАМФ. Дифузуючи в клітину, цАМФ реагує з аденілаткіназою (А-кіназа); при цьому вивільняється субодиниця С, яка каталізує фосфорилювання білка Р.Це фосфорилювання переводить білки в активну форму, і тепер вони можуть виявити свою специфічну регулюючу дію (наприклад, викликати деградацію глікогену). Ця складна регуляторна система надзвичайно ефективна, оскільки кінцевим результатом є фосфорилювання множини білків, тобто регуляторний сигнал проходить ланцюг з більшим коефіцієнтом посилення. Зовнішні медіатори, що зв'язуються з рецепторами R s та R i специфічними для кожного з них надзвичайно різноманітні. Адреналін, зв'язуючись з R s або R i бере участь у регуляції метаболізму ліпідів та глікогену, а також у посиленні скорочення серцевого м'яза та в інших реакціях (див. гл. 19). Тиреотропний гормон, активуючи R s , стимулює виділення щитовидної залози гормону тироксину, а простагландин I гальмує агрегацію кров'яних пластинок Інгібіторні ефекти, у тому числі адреналіну, опосередковані через R i виражаються у уповільненні ліполізу. Таким чином, система цАМФ-це багатофункціональна внутрішньоклітинна регуляторна система,яка може точно контролюватись позаклітинними стимуляторними та інгібіторними сигнальними речовинами.
Інозитолфосфат «ІФ з ». Внутрішньоклітинна система другого посередника – інозитолфосфату – була відкрита лише недавно (рис. 1.16). В даному випадку інгібіторний шлях відсутній, однак є подібність до системи цАМФ, в якій ефект стимуляції рецептора R переноситься на ГТФ-активований G-білок на внутрішній поверхні мембрани. На наступному етапі звичайний мембранний ліпід фосфатидилінозитол (ФІ), попередньо отримавши дві додаткові фосфатні групи, перетворюється на ФІ-дифосфат (ФІФ 2), який розщеплюється активованою фосфодіестеразою (ФДЕ) на інозитолтріфосфат(ІФз) та ліпід діацилгліцерол(ДАГ). Інозитолтрифосфат-це водорозчинний другий посередник, що дифузує в цитозоль. Він діє насамперед шляхом вивільнення Ca 2+ з ендоплазматичного ретикулуму. Ca 2+ у свою чергу діє як посередник, як було описано вище; наприклад, він активує Ca 2+ -залежну фосфокіназу, що фосфорилює ферменти. Ліпідна субодиниця ДАГ (рис. 1.16) також переносить сигнал, дифузуючи в ліпідній фазі плазматичної мембрани до розташованої на її внутрішній поверхні С-кіназі, яка активується за участю фосфатидилсерину як кофактора. Потім С-кіназа запускає фосфорилювання білків, переводячи їх у активну форму.
Внутрішньоклітинна система другого посередника ІФ також може керуватися безліччю зовнішніх медіаторів і гормонів, у тому числі ацетилхоліном, серотоніном, вазопресином і тиреотропним гормоном; як і система цАМФ, вона характеризується різноманітними внутрішньоклітинними ефектами. Можливо, ця система активується і світлом у зоровому рецепторі ока, і відіграє центральну роль фототрансдукції (див. гл. 11). Вперше в індивідуальному розвитку організму рецептор системи ІФз активується спермієм, внаслідок чого ІФз бере участь у регуляторних реакціях, що супроводжують запліднення яйцеклітини.
Системи цАМФ та ІФз-ДАГ є високоефективними біологічними підсилювачами.Вони

Рис. 1.16.Ланцюг реакцій за участю внутрішньоклітинного посередника ІФЗ (інозитолтріфосфату). Як і в системі цАМФ позаклітинний сигнал опосередкований через білокG, який у даному випадку активує фосфодіестеразу (ФДЕ). Цей фермент розщеплює фосфатидилінозиндіфосфат (ФІФ 2 ) у плазматичній мембрані до ІФз та діацилгліцеролу (ДАГ); ІФз дифундує в цитоплазму. Тут він викликає викид Са 2+ з ендоплазматичного ретикулуму; підвищення концентрації Са 2+ в цитоплазмі ([Са 2+ ] i ) активує протеїнкіназу, яка фосфорилює і, отже, активує ферменти. Інший продукт, ДАГ, залишається в мембрані та активує протеїнкіназу С (кофактор-фосфатидилсерін, ФС). Протеїнкіназа С також фосфорилює ферменти, що опосередковують специфічну дію, пов'язану зі стимуляцією зовнішнього рецептора R . Гілки ланцюга реакцій за участю ІФз та ДАГ можуть бути активовані незалежно іономіцином та форболовим ефіром відповідно (із змінами)
перетворюють реакцію між медіатором та зовнішнім мембранним рецептором у фосфорилювання множини внутрішньоклітинних білків, які потім можуть впливати на різні функції клітини. Один із суттєвих аспектів проблеми полягає в тому, що, наскільки відомо на сьогоднішній день, існують лише ці дві тісно пов'язані регуляторні системи такого типу, що використовуються численними зовнішніми посередниками для регулювання різноманітних внутрішньоклітинних процесів. Разом про те, ці регуляторні системи, зокрема і Са 2+ , тісно взаємодіють друг з одним, що дозволяє здійснювати тонку регуляцію клітинних функцій.
Підручники та керівництва
1. Alberts В., Bray D., Lewis J., Raff M., Roberts До., Watson J.D.Molecular Biology of Cell, New York and London, Garland Publishing Inc., 1983.
2. Czihak G., Longer H., Ziegler H.(Eds.). Biologie. Berlin, Heidelberg, New York, Springer, 1983.
3. Hille В. Ionic channels of excitable membranes. Sunderland, Mass., Sinauer Assoc., 1984.
4. Hoppe W., Lohmann W. Marki H., Ziegler H.(Eds.). Biophysik. Berlin, Heidelberg, New York, Springer, 1984.
5. Jungermann До., Mahler H.Biochemie. Berlin, Heidelberg, New York, Springer, 1980.
6. Kandel E. R., Schwartz-J. H.,(Eds.). Principles of neural science, New York, Amsterdam, Oxford, Elsevier, 1985.
7. Schiebler Т. H. Schmidt W.Anatomic des Menschen. Berlin, Heidelberg, New York, Tokyo, Springer, 1983.
Оригінальні статті та огляди
8. Berridge MJ.Молекулярна основа комунікації з селом, Sci. Amer, 253, 124134 (1985).
9. Berridge MJ, Irvine RF. Inositol triphosphate, a novel second messenger в cellular signal transduction. Nature, 312, 315321 (1984).
10. Bretscher M.S. Molecules of cell membrane, Sci. Amer., 253, 124-134 (1985).
11. Daut J. The living cell as an energy-transducing machine. Як мінімальний модель м'якого кардіального метаболізму, Biochem. et Biophys. Acta, 895, 41-62 (1987).
12. Hodgkin A.L., Katz В. Діяльність радіаційної кампанії на електричній діяльності гіанта аксона бризок. J. Physiol. (Lond.), 108, 37-77 (1949).
13. Hodgkin A.L., Keynes R.D. Active transport of cations in giant axons from Sepia and Лоліго, J. Physiol. (Lond.), 128, 28-42 (1955).
14. Longer P. Ionic channels with conformational substates, Biophys. J., 47, 581-590 (1985).
15. Ochs S., Worth P.M. Exoplasmic transport в normal і pathologic systems. In: Physiology and Pathology of Axons, S.G. Waxmann, Ed. New York, Raven Press, 1978.
КЛІТИНА
Епітеліальна тканина.
ВИДИ ТКАНИНЬ.
БУДОВА І ВЛАСТИВОСТІ КЛІТИНИ.
Лекція №2.
1. Будова та основні властивості клітини.
2. Поняття про тканини. Види тканин.
3. Будова та функції епітеліальної тканини.
4. Види епітелію.
Мета: знати будову та властивості клітини, види тканин. Представляти класифікацію епітелію та місцезнаходження його в організмі. Вміти відрізняти епітеліальну тканину за морфологічними ознаками з інших тканин.
1. Клітина – це елементарна жива система, основа будови, розвитку та життєдіяльності всіх тварин та рослин. Наука про клітину – цитологія (грец. сytos – клітина, logos – наука). Зоолог Т.Шванн в 1839 р. вперше сформулював клітинну теорію: клітина представляє основну одиницю будови всіх живих організмів, клітини тварин і рослин подібні за своєю будовою, поза клітиною немає життя. Клітини існують як самостійні організми (найпростіші, бактерії), і в складі багатоклітинних організмів, в яких є статеві клітини, що служать для розмноження, і клітини тіла (соматичні), різні за будовою та функціями (нервові, кісткові, секреторні і т.д.). ). Розміри клітин людини знаходяться в діапазоні від 7 мкм (лімфоцити) до 200-500 мкм (жіноча яйцеклітина, гладкі міоцити). До складу будь-якої клітини входять білки, жири, вуглеводи, нуклеїнові кислоти, АТФ, мінеральні солі та вода. З неорганічних речовин у клітині міститься найбільше води (70-80%), з органічних – білків (10-20%). Основними частинами клітини є: ядро, цитоплазма, клітинна оболонка (цитолема).
ЯДРО ЦИТОПЛАЗМУ ЦИТОЛЕМА
Нуклеоплазма – гіалоплазма
1-2 ядерця - органели
Хроматин (ендоплазматична мережа
комплекс КТольджі
клітинний центр
мітохондрії
лізосоми
спеціального призначення)
Увімкнення.
Ядро клітини знаходиться в цитоплазмі та відмежовано від неї ядерної
оболонкою – нуклеолемою. Воно є місцем зосередження генів,
основною хімічною речовиною яких є ДНК. Ядро регулює формотворчі процеси клітини та всі її життєві відправлення. Нуклеоплазма забезпечує взаємодію різних ядерних структур, ядерця беруть участь у синтезі клітинних білків та деяких ферментів, хроматин містить хромосоми з генами – носіями спадковості.
Гіалоплазма (грец. hyalos – скло) – основна плазма цитоплазми,
є справжнім внутрішнім середовищем клітини. Вона поєднує всі клітинні ультраструктури (ядро, органели, включення) та забезпечує хімічну взаємодію їх один з одним.
Органели (органоїди) – це постійні ультраструктури цитоплазми, що виконують у клітині певні функції. До них відносяться:
1) ендоплазматична мережа – система розгалужених каналів та порожнин, утворена подвійними мембранами, пов'язаними з клітинною оболонкою. На стінах каналів є дрібні тільця - рибосоми, що є центрами синтезу білка;
2) комплекс К.Гольджи, або внутрішній сітчастий апарат, - має сітки та містить вакуолі різної величини (лат. vacuum - порожній), бере участь у видільній функції клітин та утворенні лізосом;
3) клітинний центр - цитоцентр складається з кулястого щільного тіла-центросфери, всередині якого лежать 2 щільні тільця - центріолі, пов'язані між собою перемичкою. Розташовується ближче до ядра, бере участь у розподілі клітини, забезпечуючи рівномірний розподіл хромосом між дочірніми клітинами;
4) мітохондрії (грец. mitos - нитка, chondros - зерно) мають вигляд зернят, паличок, ниток. Вони здійснюється синтез АТФ.
5) лізосоми - бульбашки, заповнені ферментами, які, регулюють
обмінні процеси в клітині і мають травну (фагоцитарну) активність.
6) органели спеціального призначення: міофібрили, нейрофібрили, тонофібрили, вії, ворсинки, джгутики, що виконують специфічну функцію клітини.
Цитоплазматичні включення – це непостійні утворення у вигляді
гранул, крапель та вакуолей, що містять білки, жири, вуглеводи, пігмент.
Клітинна оболонка – цитолема, або плазмолема, покриває клітину з поверхні та відокремлює її від навколишнього середовища. Є напівпроникною і регулює надходження речовин у клітину та вихід їх із неї.
Міжклітинна речовина знаходиться між клітинами. В одних тканинах воно рідке (наприклад, у крові), а в інших складається з аморфної (безструктурної) речовини.
Будь-яка жива клітина має такі основні властивості:
1) обміном речовин, або метаболізмом (головна життєва властивість),
2) чутливістю (дратівливістю);
3) здатністю до розмноження (самовостворення);
4) здатність до зростання, тобто. збільшення розмірів та обсягу клітинних структур і самої клітини;
5) здатністю до розвитку, тобто. придбання клітиною специфічних функцій;
6) секрецією, тобто. виділенням різних речовин;
7) пересуванням (лейкоцити, гістіоцити, сперматозоїди)
8) фагоцитозом (лейкоцити, макрофаги та ін.).
2. Тканина - це система клітин, подібна за походженням), будовою та функціями. До складу тканин входять також тканинна рідина та продукти життєдіяльності клітин. Вчення про тканини називається гістологією (грец. histos - тканина, logos - вчення, наука). Відповідно до особливостей будови, функції та розвитку розрізняють такі види тканин:
1) епітеліальну, або покривну;
2) сполучну (тканини внутрішнього середовища);
3) м'язову;
4) нервову.
Особливе місце в організмі людини займає кров та лімфа – рідка тканина, що виконує дихальну, трофічну та захисну функції.
В організмі всі тканини тісно пов'язані між собою морфологічно
та функціонально. Морфологічний зв'язок обумовлена тим, що різна-
ні тканини входять до складу тих самих органів. Функціональний зв'язок
виявляється в тому, що діяльність різних тканин, що входять до складу
органів, погоджена.
Клітинні та неклітинні елементи тканин у процесі життє-
діяльності зношуються та відмирають (фізіологічна дегенерація)
та відновлюються (фізіологічна регенерація). При пошкодженні
тканин відбувається їх відновлення (репаративна регенерація).
Однак не у всіх тканин цей процес протікає однаково. Епітеліаль-
ная, сполучна, гладка м'язова тканина і клітини крові регенери-
ють добре. Поперечносмугаста м'язова тканина відновлюється
лише за певних умов. У нервовій тканині відновлюються
лише нервові волокна. Поділ нервових клітин в організмі дорослого
людину не встановлено.
3. Епітеліальна тканина (епітелій) - це тканина, що покриває поверхню шкіри, рогівку ока, а також вистилає всі порожнини організму, внутрішню поверхню порожнистих органів травної, дихальної, сечостатевої систем, що входить до складу більшості залоз організму. У зв'язку з цим розрізняють покривний та залізистий епітелій.
Покривний епітелій, будучи прикордонною тканиною, здійснює:
1) захисну функцію, оберігаючи підлягають тканини від різних зовнішніх впливів: хімічних, механічних, інфекційних.
2) обмін речовин організму з навколишнім середовищем, виконуючи функції газообміну у легенях, всмоктування у тонкому кишечнику, виділення продуктів обміну (метаболітів);
3) створення умов рухливості внутрішніх органів у серозних порожнинах: серця, легенів, кишечника тощо.
Залізистий епітелій здійснює секреторну функцію, тобто утворює і виділяє специфічні продукти - секрети, що використовуються в процесах, що протікають в організмі.
Морфологічно епітеліальна тканина відрізняється від інших тканин організму такими ознаками:
1) вона завжди займає прикордонне положення, оскільки розташовується на межі зовнішнього та внутрішнього середовищ організму;
2) вона є пластами клітин - епітеліоцитів, які мають неоднакову форму і будову в різних видах епітелію;
3) між клітинами епітелію немає міжклітинної речовини, та клітини
пов'язані один з одним за допомогою різних контактів.
4) клітини епітелію розташовані на базальній мембрані (пластинці товщиною близько 1 мкм, якою він відокремлений від сполучної тканини, що підлягає. Базальна мембрана складається з аморфної речовини і фібрилярних структур;
5) клітини епітелію мають полярність, тобто. базальні та верхівкові відділи клітин мають різну будову;
6) епітелій не містить кровоносних судин, тому харчування клітин
здійснюється шляхом дифузії поживних речовин через базальну мембрану з тканин;
7) наявність тонофібрил – нитчастих структур, що надають міцності епітеліальним клітинам.
4. Існує кілька класифікацій епітелію, в основу яких покладені різні ознаки: походження, будова, функції. З них найбільшого поширення набула морфологічна класифікація, що враховує відношення клітин до базальної мембрани та їх форму на вільній апікальній (лат. apex – вершина) частині епітеліального пласта . У цій класифікації відбито будову епітелію, що залежить від його функції.
Одношаровий плоский епітелій представлений в організмі ендотелієм та мезотелієм. Ендотелій вистилає кровоносні, лімфатичні судини, камери серця. Мезотелій покриває серозні оболонки порожнини очеревини, плеври та перикарда. Одношаровий кубічний епітелій вистилає частину ниркових канальців, протоки багатьох залоз та дрібні бронхи. Одношаровий призматичний епітелій має слизова оболонка шлунка, тонкого та товстого кишечника, матки, маткових труб, жовчного міхура, ряду проток печінки, підшлункової залози, частини.
канальців нирки. В органах, де відбуваються процеси всмоктування, епітеліальні клітини мають всмоктуючу облямівку, що складається з великої кількості мікроворсинок. Одношаровий багаторядний миготливий епітелій вистилає повітроносні шляхи: порожнину носа, носоглотку, гортань, трахею, бронхи та ін.
Багатошаровий плоский неороговіючий епітелій покриває зовні рогівку ока і слизову оболонку порожнини рота і стравоходу. Перехідний епітелій типовий для сечовідвідних органів: балій нирок, сечоводів, сечового міхура, стінки яких схильні до значного розтягування при наповненні сечею.
Екзокринні залози виділяють свій секрет у порожнині внутрішніх органів або на поверхню тіла. Вони, як правило, мають вивідні протоки. Ендокринні залози не мають проток і виділяють секрет (гормони) у кров або лімфу.
Зв'язок організму з навколишнім середовищем, з фізико-хімічної точки зору, є відкритою системою, тобто системою, де біохімічні процеси йдуть постійно. Вихідні речовини надходять із навколишнього середовища, а речовини, що утворюються також безперервно, виносяться зовні. Рівновага між швидкістю і концентрацією продуктів різноспрямованих реакцій в організмі є умовним, уявним, тому що надходження та винесення речовин не припиняються. Безперервний зв'язок із навколишнім середовищем і дозволяє розглядати живий організм як відкриту систему.
Для всіх живих клітин джерелом енергії є Сонце. Рослинні клітини вловлюють енергію сонячного світла з допомогою хлорофілу, використовуючи її реакцій асиміляції у процесі фотосинтезу. Клітини тварин, грибів, бактерій використовують сонячну енергію побічно при розщепленні органічних речовин, синтезованих земною рослиною.
Частина поживних речовин клітини розщеплюється у процесі клітинного дихання, поставляючи в такий спосіб енергію, необхідну різного роду клітинної активності. Протікає цей процес у органелах, званих мітохондріями. Мітохондрія складається з двох мембран: зовнішньої, що відокремлює органеллу від цитоплазми, та внутрішньої, що утворює численні складики. Головним продуктом дихання є АТФ. Він залишає мітохондрії і використовується як джерело енергії для багатьох хімічних реакцій у цитоплазмі та клітинній мембрані. Якщо для здійснення клітинного дихання потрібен кисень, то дихання називають аеробним, якщо ж реакції йдуть за відсутності кисню, то говорять про анаеробне дихання.
Для будь-якого виду роботи, що виконується в клітині, використовується енергія в одній формі - у формі енергії фосфатних зв'язків АТФ. АТФ – легко рухоме з'єднання. Утворення АТФ відбувається на внутрішній мембрані мітохондрій. АТФ синтезується у всіх клітинах при диханні за рахунок енергії окиснення вуглеводів, жирів та інших органічних речовин. У клітинах зелених рослин основна кількість АТФ синтезується у хлоропластах за рахунок сонячної енергії. Вони при фотосинтезі утворюється у багато разів більше АТФ, ніж у мітохондріях. Розкладається АТФ з розривом фосфорно-кисневих зв'язків та виділенням енергії. Це відбувається під дією ферменту АТФази у процесі гідролізу АТФ – приєднання води з відщепленням молекули фосфорної кислоти. В результаті АТФ перетворюється на АДФ, а якщо відщеплюються дві молекули фосфорної кислоти, то на АМФ. Реакція відщеплення кожної грам-молекули кислоти супроводжується визволенням 40 кДж. Це дуже великий вихід енергії, тому фосфорно-кисневі зв'язки АТФ називають макроергітичними (високоенергетичними).
Використання АТФ у реакціях пластичного обміну здійснюється шляхом їх поєднання з гідролізом АТФ. Молекули різних речовин заряджаються енергією шляхом приєднання звільненої при гідролізі фосфорної групи молекули АТФ, т. е. шляхом фосфорилирования.
Особливість фосфатних похідних у тому, що вони можуть залишити клітину, хоча їх «розряджені» форми вільно проходять через мембрану. Завдяки цьому фосфорильовані молекули залишаються в клітині доти, доки вони не будуть використані у відповідних реакціях.
Зворотний процес перетворення АДФ на АТФ відбувається шляхом приєднання молекули фосфорної кислоти до АДФ з виділенням води та поглинанням великої кількості енергії.
Таким чином АТФ - універсальне та безпосереднє джерело енергії для діяльності клітини. Це створює єдиний клітинний фонд енергії і уможливлює її перерозподіл та транспортування з одних ділянок клітини в інші.
Перенесення фосфатної групи відіграє у хімічних реакціях типу складання макромолекул з мономерів. Наприклад, амінокислоти можуть з'єднуватися в пептиди, лише попередньо профосфорилированными. Механічні процеси скорочення або руху, перенесення розчиненої речовини проти концентрації градієнта та інші процеси пов'язані з витрачанням енергії, запасеної в АТФ.
Процес енергетичного обміну може бути представлений в такий спосіб. Високомолекулярні органічні речовини в цитоплазмі ферментативно, шляхом гідролізу перетворюються на простіші, з яких вони складаються: білки - на амінокислоти, полі- та дисахариди - на моносахариди (+ глюкозу), жири на гліцерин і жирні кислоти. Окисні процеси відсутні, звільняється мало енергії, яка не використовується і переходить у теплову форму. Більшість клітин насамперед витрачають вуглеводи. Полісахариди (крохмаль у рослин та глікоген у тварин) гідралізуються до глюкози. Окислення глюкози відбувається у три фази: гліколіз, окисне декарбоксилювання (цикл Кребса – цикл лимонної кислоти) та окисне фосфорилювання (дихальний ланцюг). Гліколіз, внаслідок якого одна молекула глюкози розщеплюється на дві молекули піровиноградної кислоти з виділенням двох молекул АТФ, протікає у цитоплазмі. За відсутності кисню піровиноградна кислота перетворюється або на етанол (бродіння), або молочну кислоту (анаеробне дихання).
Якщо гліколіз здійснюється у клітинах тварин, шестивуглецева молекула глюкози розпадається на дві молекули молочної кислоти. Процес цей багатоступінчастий. Його послідовно здійснюють 13 ферментів. При спиртовому бродінні з молекули глюкози утворюється дві молекули етанолу та дві молекули СО2.
Гліколіз - фаза, загальна для анаеробного та аеробного дихання, дві інші здійснюються лише в аеробних умовах. Процес безкисневого окислення, у якому виділяється і використовується лише частина енергії метаболітів, для анаеробних організмів є кінцевим. У присутності ж кисню піровиноградна кислота переходить у мітохондрії, де в результаті цілого ряду послідовних реакцій вона повністю окислюється аеробним шляхом до Н2O та СО2 з одночасним фосфорилуванням АДФ до АТФ. При цьому дві молекули АТФ дає гліколіз, дві – цикл Кребса, 34 – дихальний ланцюг. Чистий вихід при повному окисленні однієї молекули глюкози до Н2О та СО2 становить 38 молекул.
Таким чином, у аеробних організмів остаточний розпад органічних речовин здійснюється шляхом окиснення їх киснем повітря до простих неорганічних: СО2 та Н2О. Процес цей протікає на кристалах мітохондрій. При цьому виділяється максимальна кількість вільної енергії, значна частина якої резервується молекулами АТФ. Легко бачити, що аеробне окиснення найбільше забезпечує клітину вільної енергією.
В результаті катаболізму в клітині накопичуються багаті енергією молекули АТФ, а у зовнішнє середовище виділяється СО2 та надмірна кількість води.
Молекули цукрів, які не потрібні для дихання, можуть запасатися в клітці. Надлишкові ліпіди або розщеплюються, після чого продукти їх розщеплення надходять у мітохондрії як субстрат для дихання, або відкладаються про запас у цитоплазмі у вигляді жирових крапель. З амінокислот, що надходять у клітину, будуються білки. Синтез білка відбувається у органелах, званих рибосомами. Кожна рибосома складається з двох субчастинок - великої та малої: до складу обох субчастинок входять білкові молекули та молекули РНК.
Рибосоми часто бувають прикріплені до особливої системи мембран, що складається з цистерн та бульбашок, - до так званого ендоплазматичного ретикулуму (ЕР); в клітинах, що виробляють багато білка, ендоплазматичний ретикулум часто розвинений дуже добре і весь усіяний рибосомами. Деякі ферменти ефективні лише тому випадку, якщо вони прикріплені до мембрани. Тут перебуває більшість ферментів, що у синтезі ліпідів. Таким чином, ендоплазматичний ретикулум - це своєрідний робочий стіл клітини.
Крім того, ЕР ділить цитоплазму на окремі відсіки, або компартменти, тобто роз'єднує різні хімічні процеси, що одночасно протікають у цитоплазмі, і тим самим знижує ймовірність того, що ці процеси заважатимуть один одному.
Часто утворені цією клітиною продукти використовуються поза цією клітиною. У подібних випадках синтезовані на рибосомах білки проходять через мембрани ендоплазматичного ретикулуму і упаковуються в мембранні бульбашки, що формуються навколо них, які потім відшнуровуються від ЕР. Ці бульбашки, уплощаясь і укладаючись один на одного, як млинці в стопці, утворюють характерну структуру, яка називається комплексом Гольджі, або апаратом Гольджі. Під час свого перебування в апараті Гольджі, білки зазнають певних змін. Коли їм настає час залишити клітину, мембранні бульбашки зливаються з клітинної мембраною і спорожняються, виливаючи свій вміст назовні, т. е. секреція відбувається шляхом екзоцитозу.
В апараті Гольджі утворюються також лізосоми – мембранні мішечки, що містять травні ферменти. З'ясування того, як клітина виробляє, упаковує та експортує деякі білки, а також як вона «дізнається», які білки їй слід зберігати для себе, становить один із найцікавіших розділів сучасної цитології.
Мембрани будь-якої клітини безперервно переміщуються і видозмінюються. Мембрани ЕР повільно переміщуються клітиною. Окремі ділянки цих мембран відокремлюються і утворюють бульбашки, які на якийсь час стають частиною апарату Гольджі, а потім, в процесі екзоцитозу, зливаються з клітинною мембраною.
Пізніше мембранний матеріал повертається до цитоплазми, де він використовується знову.
Обмін речовин, що надходять всередину клітини або виділяються назовні, а також обмін різними сигналами з мікро-і макрооточенням відбувається через зовнішню мембрану клітини. Як відомо, клітинна мембрана є ліпідним біслоєм, в який вбудовані різні білкові молекули, що виконують роль спеціалізованих рецепторів, іонних каналів, пристроїв, які активно переносять або видаляють різні хімічні речовини, міжклітинних контактів і т.п. асиметрично: зовнішня поверхня складається із сфінгомієліну та фосфатидилхоліну, внутрішня - з фосфатидилсерину та фосфатидилетаноламіну. Підтримка такої асиметрії потребує витрат енергії. Тому у разі пошкодження клітини, її інфікування, енергетичного голодування відбувається збагачення зовнішньої поверхні мембрани невластивими їй фосфоліпідами, що стає сигналом для інших клітин та ферментів про пошкодження клітини з відповідною реакцією на це. Найбільшу роль грає розчинна форма фосфоліпази A2, яка розщеплює арахідонову кислоту та створює лізоформи з вищеназваних фосфоліпідів. Арахидонова кислота є лімітуючою ланкою для створення таких медіаторів запалення, як ейкозаноїди, а до лізоформ в мембрані приєднуються захисні молекули - пентраксини (С-реактивний протеїн (СРП), попередники амілоїдних білків) - з подальшою активацією системи.
Будова мембрани сприяє збереженню особливостей внутрішнього середовища клітини, її відмінностей від зовнішнього середовища. Це забезпечується вибірковою проникністю клітинної мембрани, існуванням у ній механізмів активного транспорту. Їх порушення в результаті прямого пошкодження, наприклад, тетродотоксином, уабаїном, тетраетиламмонием, або у разі недостатнього енергетичного забезпечення відповідних "насосів" призводить до порушення електролітного складу клітини, зміни в ній метаболізму, порушення специфічних функцій - скорочення, проведення імпульсу збудження і т.п. Порушення клітинних іонних каналів (кальцієвих, натрієвих, калієвих та хлорних) у людини також може бути генетично обумовлено мутацією генів, які відповідають за структуру цих каналів. Так звані каналопатії спричиняють спадкові хвороби нервової, м'язової, травної систем. Надмірне надходження всередину клітини води може призвести до її розриву – цитолізу – внаслідок перфорації мембрани при активації комплементу або атаки цитотоксичних лімфоцитів та натуральних кілерів.
У клітинну мембрану вбудовано безліч рецепторів – структур, які у разі поєднання з відповідними специфічними сигнальними молекулами (лігандами) передають сигнал усередину клітини. Це відбувається за допомогою різних регуляторних каскадів, що складаються з ферментативно-активних молекул, які послідовно активуються і в кінцевому підсумку сприяють реалізації різних клітинних програм, таких як ріст і проліферація, диференціювання, рухливість, старіння, загибель клітини. Регуляторні каскади досить численні, та їх кількість остаточно ще визначено. Система рецепторів та об'єднаних з ними регуляторних каскадів існує і всередині клітини; вони створюють певну регуляторну мережу з пунктами концентрації, розподілу та вибору подальшого шляху сигналу залежно від функціонального стану клітини, етапу її розвитку, одночасної дії сигналів з інших рецепторів. Результатом цього може бути гальмування або посилення сигналу, направлення його іншим регуляторним шляхом. Як рецепторний апарат, так і шляхи передачі сигналу через регуляторні каскади, наприклад до ядра, можуть порушуватися в результаті генетичного дефекту, який виникає як вроджена вада на рівні організму або внаслідок соматичної мутації в певному типі клітин. Зазначені механізми можуть ушкоджуватися інфекційними агентами, токсинами, і навіть змінюватися у процесі старіння. Завершальним етапом цього може бути порушення функцій клітини, процесів її проліферації та диференціювання.
На поверхні клітин також розташовані молекули, що відіграють важливу роль у процесах міжклітинної взаємодії. До них можуть відноситися білки клітинної адгезії, антигени тканинної сумісності, тканинноспецифічні, диференціюючі антигени тощо. Зміни у складі цих молекул зумовлюють порушення міжклітинних взаємодій і можуть стати причиною включення відповідних механізмів елімінації таких клітин, адже вони представляють резервуар інфекції, особливо вірусної, чи як потенційні ініціатори пухлинного зростання.
Порушення енергетичного забезпечення клітини
Джерелом енергії у клітині служать продукти харчування, після розщеплення яких до кінцевих речовин виділяється енергія. Головним місцем утворення енергії є мітохондрії, в яких речовини окислюються за допомогою ферментів дихального ланцюга. Окислення - основний постачальник енергії, оскільки в результаті гліколізу з такої кількості субстратів окислення (глюкози) вивільняється, порівняно з окисленням, не більше 5 % енергії. Близько 60 % енергії, що вивільняється при окисненні, акумулюється шляхом окисного фосфорилювання в макроергічних фосфатах (АТФ, креатинфосфат), решта її розсіюється у вигляді тепла. В подальшому макроергічні фосфати використовуються клітиною для таких процесів, як робота насосів, синтез, розподіл, рух, секреція та ін. , третій – механізм використання енергії.
Порушення електронного транспорту в дихальному ланцюгу мітохондрій або роз'єднання окиснення та фосфорилювання АДФ при втраті протонного потенціалу - рушійної сили генерації АТФ, призводить до ослаблення окисного фосфорилювання таким чином, що велика частина енергії розсіюється у вигляді тепла і зменшується кількість макроерг. Роз'єднання окислення та фосфорилювання під впливом адреналіну використовується клітинами гомойотермних організмів для підвищення теплопродукції за підтримки постійної температури тіла під час охолодження або її підвищення при лихоманці. Значні зміни у структурі мітохондрій та енергетичному метаболізмі спостерігаються при тиреотоксикозі. Ці зміни спочатку оборотні, але після певної риси стають незворотними: мітохондрії фрагментуються, розпадаються або набухають, втрачають кристи, перетворюючись на вакуолі, і зрештою накопичують такі речовини, як гіалін, феритин, кальцій, ліпофусцин. У хворих на цингу відбувається злиття мітохондрій з утворенням хондріосфер, можливо, внаслідок пошкодження мембран пероксидними сполуками. Значні пошкодження мітохондрій виникають під дією іонізуючого випромінювання, в ході перетворення нормальної клітини на злоякісну.
Мітохондрії є потужним депо іонів кальцію, де його концентрація на кілька порядків перевищує таку в цитоплазмі. При пошкодженні мітохондрій кальцій виходить у цитоплазму, зумовлюючи активізацію протеїназ із пошкодженням внутрішньоклітинних структур та порушенням функцій відповідної клітини, наприклад, кальцієві контрактури або навіть “кальцієву смерть” у нейронах. Внаслідок порушення функціональної здатності мітохондрій різко збільшується утворення вільнорадикальних пероксидних сполук, які мають дуже високу реакційну здатність і тому ушкоджують важливі компоненти клітини – нуклеїнові кислоти, білки та ліпіди. Це спостерігається при так званому оксидаційному стресі і може мати негативні наслідки для існування клітини. Так, пошкодження зовнішньої мембрани мітохондрії супроводжується виходом у цитоплазму речовин, що містяться в міжмембранному просторі, насамперед цитохрому С та деяких інших БАВ, які запускають ланцюгові реакції, що є причиною запрограмованої загибелі клітини – апоптозу. Пошкоджуючи ДНК мітохондрій, вільнорадикальні реакції спотворюють генетичну інформацію, необхідну для утворення деяких ферментів дихального ланцюга, які продукуються саме в мітохондріях. Це призводить до ще більшого порушення окисних процесів. У цілому нині власний генетичний апарат мітохондрій проти генетичним апаратом ядра гірше захищений від шкідливих впливів, здатних змінювати закодовану у ньому генетичну інформацію. Як результат – виникає порушення функцій мітохондрій протягом життя, наприклад, у процесі старіння, при злоякісній трансформації клітини, а також на тлі спадкових мітохондріальних хвороб, пов'язаних із мутацією ДНК мітохондрій у яйцеклітині. В даний час описано понад 50 мітохондріальних мутацій, що викликають спадкові дегенеративні хвороби нервової та м'язової систем. Вони передаються дитині виключно від матері, тому що мітохондрії сперматозоїда не входять до складу зиготи і, відповідно, нового організму.
Порушення збереження та передачі генетичної інформації
Ядро клітини містить більшу частину генетичної інформації, і тим самим забезпечує її нормальне функціонування. За допомогою вибіркової експресії генів воно координує роботу клітини в інтерфазі, зберігає генетичну інформацію, відтворює та передає генетичний матеріал у процесі розподілу клітин. У ядрі відбуваються реплікація ДНК та транскрипція РНК. Різні патогенні фактори, такі як ультрафіолетове та іонізуюче випромінювання, вільнорадикальне окиснення, хімічні речовини, віруси, здатні пошкоджувати ДНК. Підраховано, що кожна клітина теплокровної тварини за 1 добу. втрачає понад 10 000 підстав. Сюди слід додати порушення під час копіювання під час поділу. За збереження цих пошкоджень клітина була б здатна вижити. Захист полягає у існуванні потужних систем репарації, таких як ультрафіолетова ендонуклеаза, система репаративної реплікації та рекомбінаційного відновлення, які заміняють порушення ДНК. Генетичні дефекти в репаративних системах викликають розвиток хвороб, зумовлених підвищеною чутливістю до факторів, що ушкоджують ДНК. Це пігментна ксеродерма, також деякі синдроми прискореного старіння, що супроводжуються підвищеною схильністю до виникнення злоякісних пухлин.
Система регулювання процесів реплікації ДНК, транскрипції інформаційною РНК (іРНК), трансляції генетичної інформації з нуклеїнових кислот у структуру білків досить складна та багаторівнева. Крім регуляторних каскадів, що запускають дію факгор транскрипції загальною кількістю понад 3000, які активують певні гени, існує ще багаторівнева регуляторна система, опосередкована малими молекулами РНК (інтерферуючі РНК; РНКи). Геном людини, що складається приблизно з 3 млрд пуринових та піримідинових основ, містить лише 2% структурних генів, які відповідають за синтез білків. Інші забезпечують синтез регуляторних РНК, які одночасно з факторами транскрипції активують або блокують роботу структурних генів на рівні ДНК у хромосомах або впливають на процеси трансляції матричної РНК (мРНК) під час утворення молекули поліпептиду в цитоплазмі. Порушення генетичної інформації може відбуватися на рівні структурних генів, і регуляторної частини ДНК з відповідними проявами як різноманітних спадкових захворювань.
Останнім часом велику увагу привертають зміни у генетичному матеріалі, що відбуваються у процесі індивідуального розвитку організму та пов'язані з гальмуванням або активацією певних ділянок ДНК та хромосом внаслідок їх метилювання, ацетилювання та фосфорилювання. Ці зміни зберігаються тривалий час, іноді - протягом усього життя організму від ембріогенезу до старості, і отримали назву епігеномної спадковості.
Розмноженню клітин із зміненою генетичною інформацією перешкоджають також системи (чинники) контролю мітотичного циклу. Вони взаємодіють з циклінзалежними протеїнкіназами та їх каталітичними субодиницями - циклінами - і блокують проходження клітиною повного мітотичного циклу, зупиняючи поділ на межі між пресинтетичною та синтетичною фазами (блок G1/S) аж до завершення репарації ДНК, а у разі її неможливості - клітини. До таких факторів відноситься ген р53, мутація якого спричиняє втрату контролю над проліферацією трансформованих клітин; вона спостерігається майже 50 % випадків раку в людини. Другий контрольний пункт проходження мітотичного циклу знаходиться на межі G2/M. Тут контролюється правильність розподілу хромосомного матеріалу між дочірніми клітинами в мітозі або мейозі за допомогою комплексу механізмів, що контролюють клітинне веретено, центр та центроміри (кінетохори). Неефективність цих механізмів призводить до порушення розподілу хромосом або їх частин, що проявляється відсутністю будь-якої хромосоми в одній з дочірніх клітин (анеуплоїдія), наявністю зайвої хромосоми (поліплоїдія), відривом частини хромосоми (делеція) та перенесенням її на іншу хромосому . Такі процеси дуже часто спостерігаються при розмноженні злоякісно перероджених та трансформованих клітин. Якщо це відбувається під час мейозу зі статевими клітинами, то призводить або до загибелі плода на ранньому етапі ембріонального розвитку, або до народження організму з хромосомною хворобою.
Неконтрольоване розмноження клітин під час зростання пухлин виникає як результат мутації в генах, які контролюють проліферацію клітин та отримали назву онкогенів. Серед більш ніж 70 відомих в даний час онкогенів більша частина відноситься до компонентів регуляції росту клітини, частина представлена факторами транскрипції, що регулюють активність генів, а також факторами, які гальмують розподіл та зростання клітин. Ще одним фактором, що обмежує надмірну експансію (поширення) проліферуючих клітин, є скорочення кінців хромосом - теломер, які не здатні в результаті суто стеричного взаємодії повністю реплікуватися, тому після кожного поділу клітини теломери коротшають на певну частину підстав. Таким чином, клітини дорослого організму, що проліферують, після певної кількості поділів (зазвичай від 20 до 100 залежно від виду організму та його віку) вичерпують довжину теломери і подальша реплікація хромосом припиняється. Це явище не виникає у сперматозогенному епітелії, ентероцитах та ембріональних клітинах завдяки наявності ферменту теломерази, що відновлює довжину теломер після кожного поділу. У більшості клітин дорослих організмів теломераза заблокована, але, на жаль, вона активована у клітинах пухлин.
Зв'язок між ядром та цитоплазмою, транспорт речовин в обох напрямках здійснюються через пори в ядерній мембрані за участю спеціальних транспортних систем із споживанням енергії. Таким чином транспортуються до ядра енергетичні та пластичні речовини, сигнальні молекули (чинники транскрипції). Зворотний потік виносить у цитоплазму молекули іРНК та транспортної РНК (тРНК), рибосоми, необхідні для синтезу білка у клітині. Цей шлях транспорту речовин притаманний і вірусам, зокрема таким, як ВІЛ. Вони переносять свій генетичний матеріал у ядро клітини-господаря з подальшим включенням його в геном господаря та перенесенням новоутвореної вірусної РНК у цитоплазму для подальшого синтезу білків нових вірусних частинок.
Порушення процесів синтезу
p align="justify"> Процеси синтезу білків відбуваються в цистернах ендоплазматичної мережі, тісно пов'язаних з порами в ядерній мембрані, через які в ендоплазматичну мережу надходять рибосоми, тРНК та іРНК. Тут здійснюється синтез поліпептидних ланцюжків, які надалі набувають свого остаточного вигляду в агранулярній ендоплазматичній мережі та пластинчастому комплексі (комплексі Гольджі), де піддаються трансляційній модифікації та з'єднанню з молекулами вуглеводів і ліпідів. Новоутворені білкові молекули не залишаються на місці синтезу, а за допомогою складного регульованого процесу, що зветься протеїнкінезису, активно переносяться до тієї ізольованої частини клітини, де вони виконуватимуть призначену їм функцію При цьому дуже важливим етапом є структурування перенесеної молекули відповідну просторову конфігурацію, здатну виконувати властиву їй функцію. Таке структурування відбувається за допомогою спеціальних ферментів або на матриці спеціалізованих білкових молекул - шаперонів, які допомагають білковій молекулі, новоствореної або зміненої внаслідок зовнішнього впливу, набути правильної тривимірної структури. У разі несприятливого впливу на клітину, коли виникає можливість порушення структури білкових молекул (наприклад, при підвищенні температури тіла, інфекційному процесі, інтоксикації) концентрація шаперонів у клітині різко збільшується. Тому такі молекули отримали ще назву стрес-білків, або білків теплового шоку. Порушення структурування білкової молекули призводить до утворення хімічно інертних конгломератів, які відкладаються в клітині або поза нею при амілоїдозі, хворобі Альцгеймера та ін. Іноді матрицею може служити попередньо структурована аналогічна молекула, і в даному випадку, якщо первинне структурування відбулося неправильно будуть дефектними. Ця ситуація виникає при так званих пріонових хворобах (скріпи у овець, сказ корів, куру, хвороба Крейтцфельдта-Якоба у людини), коли дефект одного з мембранних білків нервової клітини обумовлює подальше скупчення інертних мас усередині клітини та порушення її життєдіяльності.
Порушення процесів синтезу в клітині може відбуватися на різних його етапах: транскрипція РНК у ядрі, трансляція поліпептидів у рибосомах, посттрансляційна модифікація, гіперметилювання та глікозилювання бежевої молекули, транспорт та розподіл білків у клітині та виведення їх назовні. При цьому можна спостерігати збільшення або зменшення кількості рибосом, розпад полірибосом, розширення цистерн гранулярної ендоплазматичної мережі, втрату рибосом, утворення везикул і вакуолей. Так, при отруєнні блідою поганкою ушкоджується фермент РНК-полімераза, що порушує транскрипцію. Дифтерійний токсин, інактивуючи фактор елонгації, порушує процеси трансляції, зумовлюючи ушкодження міокарда. Причиною порушення синтезу деяких специфічних білкових молекул можуть бути інфекційні агенти. Наприклад, герпесвіруси гальмують синтез та експресію молекул антигенів ГКГС, що дозволяє їм частково уникнути імунного контролю, бацили чуми – синтез медіаторів гострого запалення. Поява незвичайних білків може призупиняти їх подальший розпад і призводити до накопичення інертного або токсичного матеріалу. Цьому певною мірою може сприяти порушення процесів розпаду.
Порушення процесів розпаду
Поруч із синтезом білка у клітині безупинно відбувається його розпад. У нормальних умовах це має важливе регуляторне та формотворне значення, наприклад, під час активації неактивних форм ферментів, білкових гормонів, білків мітотичного циклу. Нормальне зростання та розвиток клітини потребують тонко контрольованого балансу між синтезом та деградацією білків та органел. Однак у процесі синтезу білків внаслідок помилок у роботі синтезуючого апарату, аномального структурування білкової молекули, її пошкодження хімічними та бактеріальними агентами постійно утворюється досить багато дефектних молекул. За деякими оцінками, їхня частка становить близько третини всіх синтезованих білків.
Клітини ссавців мають кілька головних шляхів руйнування білків:через лізосомальні протеази (пентидгідролази), кальційзалежні протеїнази (ендопептидази) та систему протеасом. Крім того, є ще й спеціалізовані протеїнази, наприклад, каспази. Основною органелою, де відбувається деградація речовин в еукаріотичних клітинах, є лізосома, що містить численні гідролітичні ферменти. Внаслідок процесів ендоцитозу та різних типів аутофагії в лізосомах та фаголізосомах руйнуються як дефектні білкові молекули, так і цілі органели: пошкоджені мітохондрії, ділянки плазматичної мембрани, деякі екстрацелюлярні білки, вміст секреторних гранул.
Важливим механізмом деградації білків є протеасома - мультиката-літична протеїназна структура складної будови, що локалізується в цитозолі, ядрі, ендоплазматичній мережі та на мембрані клітини. Ця ферментна система відповідає за руйнування пошкоджених білків, а також здорових білків, які мають бути видалені для нормального функціонування клітини. При цьому білки, що підлягають руйнуванню, заздалегідь з'єднуються зі специфічним поліпептидом убіквітіном. Однак частково в протеасомах можуть руйнуватися і неубіквітовані білки. Розпад білкової молекули в протеасомах до коротких поліпептидів (процесинг) з подальшою їх презентацією разом з молекулами ГКГС І типу є важливою ланкою у здійсненні імунного контролю антигенного гомеостазу організму. При ослабленні функції протеасом відбувається акумуляція пошкоджених та непотрібних білків, що супроводжує старіння клітини. Порушення деградації циклінзалежних білків призводить до порушення клітинного поділу, деградації секреторних білків – до розвитку цистофіброзу. І навпаки, підвищення функції протеасом супроводжує виснаження організму (СНІД, рак).
При генетично обумовлених порушеннях деградації білків організм нежиттєздатний та гине на ранніх стадіях ембріогенезу. Якщо ж порушується розпад жирів чи вуглеводів, то виникають хвороби накопичення (тезаурісмози). При цьому всередині клітини накопичується надмірна кількість певних речовин або продуктів їхнього неповного розпаду - ліпідів, полісахаридів, що суттєво ушкоджує функцію клітини. Найчастіше це спостерігається в епітеліоцитах печінки (гепатоцитах), нейронах, фібробластах та макрофагоцитах.
Набуті порушення процесів розпаду речовин можуть виникати як результат патологічних процесів (наприклад, білкової, жирової, вуглеводної та пігментної дистрофії) та супроводжуватися утворенням незвичних речовин. Порушення в системі лізосомного протеолізу призводять до зниження адаптації при голодуванні або підвищеному навантаженні, до виникнення деяких ендокринних дисфункцій – зниження рівня інсуліну, тиреоглобуліну, цитокінів та їх рецепторів. Порушення деградації білків уповільнюють швидкість загоєння ран, зумовлюють розвиток атеросклерозу, впливають імунну відповідь. При гіпоксії, зміні внутрішньоклітинного pH, променевому ураженні, що характеризуються посиленою пероксидацією мембранних ліпідів, а також під впливом лізосомотропних речовин - ендотоксин бактерій. що викликає руйнування структур клітини та її загибель.